People will forgive you for being wrong, but they will never forgive you for being right – especially if events prove you right while proving them wrong. Thomas Sowell
A 2026 primate study investigated effects of vitamin C:
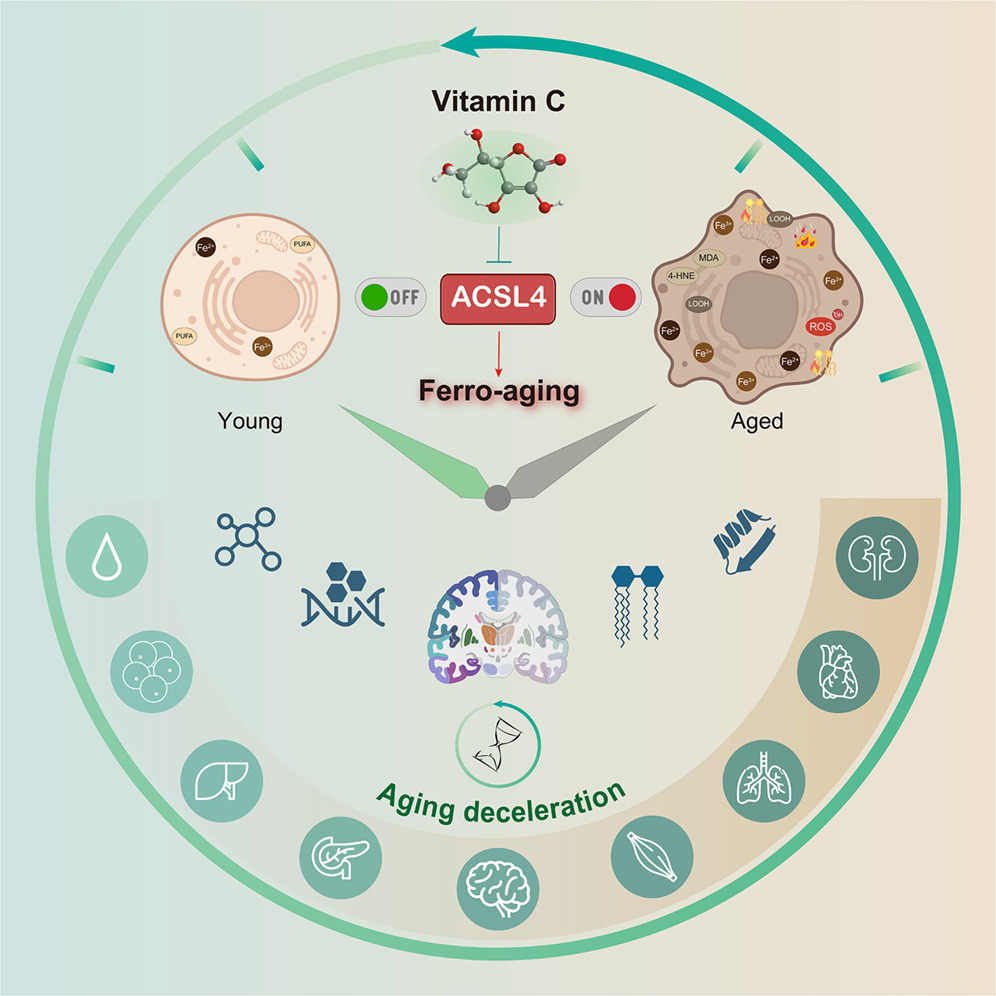
“Here, we define a conserved iron-lipid axis driving primate aging, termed ‘ferro-aging.’ Multi-tissue profiling in humans and non-human primates reveals age-progressive iron accumulation, fueling chronic lipid peroxidation orchestrated by acyl-coenzyme A (CoA) synthetase long-chain family member 4 (ACSL 4). Distinct from acute ferroptosis, this ACSL4-mediated process promotes cellular senescence and systemic functional decline.
We identify vitamin C (VC) as a direct inhibitor of ACSL4. Long-term VC administration in aged monkeys for over 40 months potently reduces ferro-aging signatures across tissues, attenuates multi-organ pathology, and improves neurological and metabolic functions. Multi-omic aging clocks indicate the VC-mediated reversal of biological age.
Despite decades of interest in oxidative stress, largely sparked by the free radical theory of aging, efforts to modulate it broadly with antioxidants have yielded inconsistent or neutral outcomes, highlighting the theory’s limitations and underscoring the need to identify more specific, upstream drivers. A critical challenge remains: determining whether the iron-lipid axis constitutes a core upstream driver of aging in primates and, if so, whether it is therapeutically targetable.
In this study, we bridge these gaps. We define an iron-triggered, ACSL4-governed, lipid peroxidation-driven program that escalates with age across diverse cell types and multiple organs in non-human primates.
VC treatment dose-dependently increased Nrf2 phosphorylation and activation. VC orchestrates a dual-defense strategy against ferro-aging: it directly suppresses the pro-aging lipid peroxidation driver ACSL4, while in parallel, it bolsters the cell’s intrinsic antioxidant capacity via Nrf2 pathway activation.
Middle-aged cynomolgus monkeys (12–16 years old, approximating human 40–50 years) received daily oral VC (30 mg/kg group) or a control treatment for 40 months under standardized conditions.
Structural MRI analysis demonstrated that VC intervention counteracted age-related brain atrophy. Using general linear mixed models, we found that VC restored cortical surface area in the frontal lobes of aged monkeys. Regional analysis identified enlargement in four regions of the orbital frontal cortex, an area critical for adaptivebehavior.
Diffusion MRI-based connectomics revealed that, compared with young animals, aged monkeys exhibited reduced structural connectivity in 18 brain regions. VC treatment restored connectivity in 9 of these regions, which were predominantly located in the posterior parietal cortex, a hub for spatial awareness and decision-making.
VC exerted robust neuroprotective effects. It attenuated heterochromatin loss (increased H3K9me3) in the prefrontal cortex and hippocampus and reduced abnormal protein aggregates, including cytosolic aggresomes and Aβ. Additionally, VC lowered the abundance of activated microglia and astrocytes and suppressed expression of the innate immune sensor cGAS in the hippocampus.
VC supplementation reduced the estimated biological age across multiple organs. At the epigenetic level, VC lowered DNA methylation age in several tissues, including brain, brown adipose tissue, muscle, skin, aorta, and kidney. In the hippocampus, the most substantial reductions in biological age occurred in microglia, oligodendroglia, and oligodendrocyte precursor cells. In the pancreas, alpha cells, beta cells, and ductal cells showed the greatest rejuvenation.
In summary, chronic VC supplementation inhibits the ferro-aging pathway, reduces multidimensional biological age across primate organs, and ameliorates a spectrum of functional declines in nervous and metabolic systems. Our work establishes ACSL4 inhibition as a promising and translationally relevant therapeutic strategy for mitigating aging-related decline.
A long-term, 40-month intervention study in aged non-human primates is a highly translational model given their shared inability with humans to synthesize VC endogenously. The finding that a single, safe nutrient can reverse multidimensional aging clocks in a primate has profound implications for translational longevity medicine.”
“For humans (who, like macaques, cannot synthesize vitamin C), the Recommended Dietary Allowance (RDA) is 75–90 mg/day for adults (~1–1.5 mg/kg for a 60–70 kg person) to prevent deficiency. Upper safe intake levels are much higher: up to 2,000 mg/day (Tolerable Upper Intake Level) is considered safe for most adults, with no established adverse effects at that level from food/supplements.
Treated monkeys represent advanced aging stages (likely equivalent to human 50s–70s+ based on ‘aged’ designation and long-term intervention effects), extending the prior 12–16-year monkey range (human ~35–55) to broader anti-aging applications. While human trials are needed, the primate evidence (long-duration, systemic benefits) strengthens the case for high-dose, sustained vitamin C as a strategy against ferro-aging in humans. It elevates vitamin C from a nutrient to a targeted anti-aging compound in primates.”
Coincidentally, I started taking extra vitamin C separately from other supplements in the form of liposomal 1 gram twice daily this past winter. Can’t say that it had any effects on my intended target, avoiding sniffles and sneezing, as allergy season kicked off in early February. With this study’s findings, I’ll continue.
Here are five 2025 human ergothioneine studies, starting with a clinical trial of healthy older adults:
“In this 16-week randomized, double-blind, placebo-controlled trial, 147 adults aged 55–79 with subjective memory complaints received ergothioneine (10 mg or 25 mg/day ErgoActive®) or placebo. Across all the groups, approximately 73% of participants in each group were female, with a median age of 69 years.
The primary outcome was the change in composite memory. Secondary outcomes included other cognitive domains, subjective memory and sleep quality, and blood biomarkers. At baseline, participants showed slightly above-average cognitive function (neurocognitive index median = 105), with plasma ergothioneine levels of median = 1154 nM.
Although not synthesized in the human body, ergothioneine is efficiently absorbed via the OCTN1 transporter (also known as the ergothioneine transporter, or ETT), which is expressed in many tissues, including the intestine, red blood cells, kidneys, bone marrow, immune cells, skin, and brain. This transporter enables ergothioneine to accumulate in high concentrations in organs vulnerable to oxidative stress and inflammation. Ergothioneine has multiple cellular protective functions, including scavenging reactive oxygen species, chelating redox-active metals, suppressing pro-inflammatory signaling, and protecting mitochondrial function.
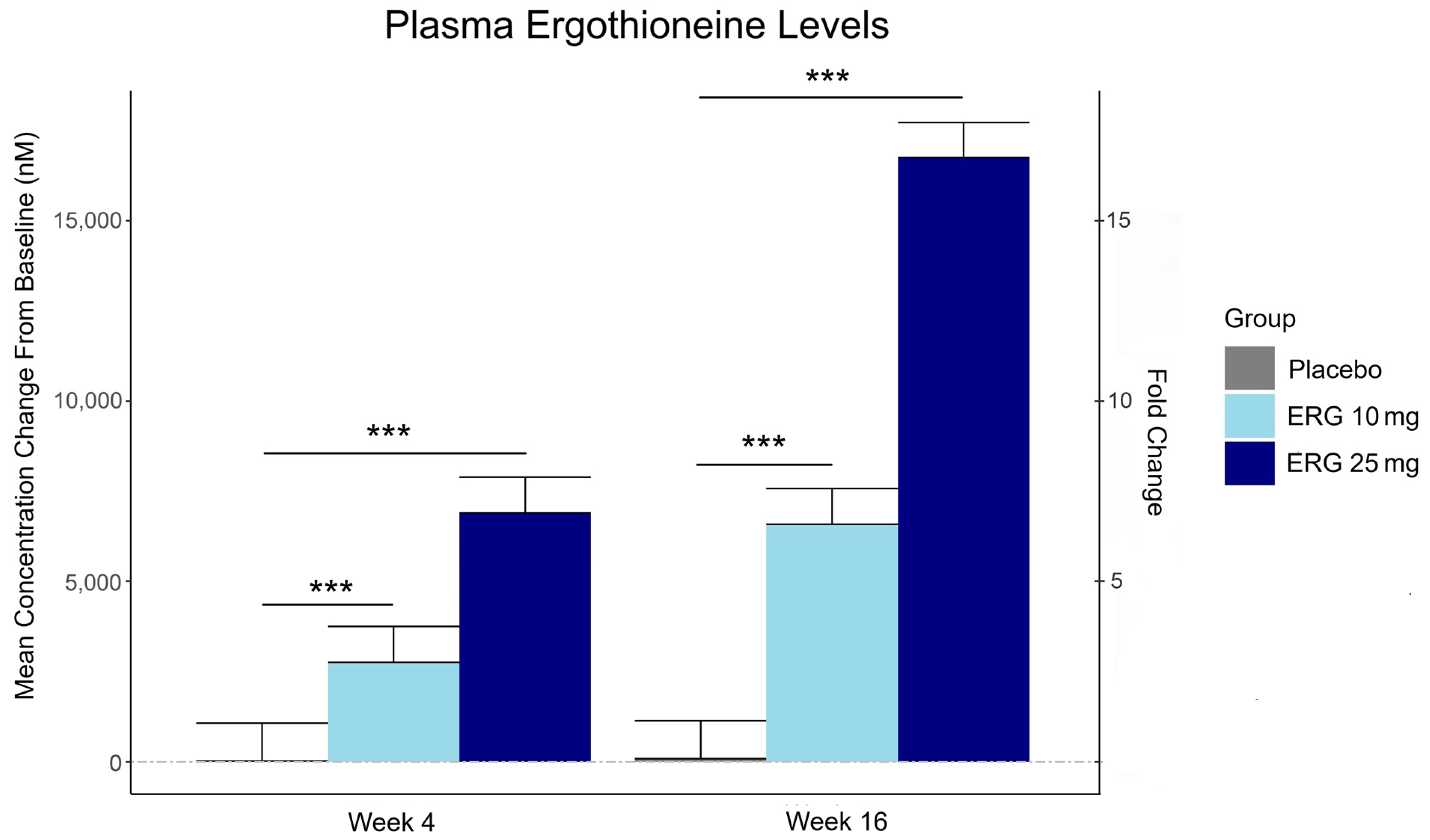
Plasma ergothioneine increased by ~3- and ~6-fold for 10 mg, and ~6- and ~16-fold for 25 mg, at weeks 4 and 16, respectively.
While the primary outcome, composite memory, showed early improvement in the 25 mg group compared to baseline, this effect was not sustained and did not differ from placebo. Reaction time showed a significant treatment-by-time interaction favoring ergothioneine, yet the between-group differences were not significant, suggesting that any potential benefits were modest and require validation in larger or longer studies.
Other cognitive effects observed were primarily within-group and not consistently dose-responsive, highlighting the challenge of detecting objective cognitive changes over a relatively short study duration in high-functioning healthy populations. However, positive effects of ergothioneine supplementation were observed on subjective measures of prospective memory and sleep initiation that were not seen in the placebo group.
This trial adds to the growing body of evidence supporting the favorable safety profile of ergothioneine. No adverse events attributable to ergothioneine were reported. Additionally, we observed potential hepatoprotective effects, with significant reductions in the plasma AST and ALT levels, particularly among males in the ERG 25 mg group.”
https://www.mdpi.com/1661-3821/5/3/15 “The Effect of Ergothioneine Supplementation on Cognitive Function, Memory, and Sleep in Older Adults with Subjective Memory Complaints: A Randomized Placebo-Controlled Trial”
The third graphic for Ergothioneine dosing, Part 2 showed a human study where a 25 mg dosing stopped after Day 7, but the plasma ergothioneine level stayed significantly higher than baseline through Day 35.
The second graphic for Ergothioneine dosing, Part 2 was a male mouse experiment where plasma ergothioneine levels of a human equivalent 22 mg to 28 mg daily dose kept rising through 92 weeks.
This trial couldn’t explain the desirability of a 25 mg daily dose that was likely (per the second and third graphics for Ergothioneine dosing, Part 2) to sustain the subjects’ increased plasma ergothioneine levels well after the trial ended at Week 16. What effects can be expected from a sustained plasma ergothioneine level that’s 16 times higher than the subjects’ initial levels? Were these 16-fold sustained plasma ergothioneine levels better or worse than the 6-fold increases in the 10 mg group, both of which were likely to continue past the trial’s end?
A representative of the trial’s sponsoring company talked a little more about the trial in this interview:
Another clinical trial investigated ergothioneine’s effects on skin:
“We conducted an 8-week, randomized, double-blind, placebo-controlled clinical trial to evaluate effects of daily oral supplementation with 30 mg of ergothioneine (DR.ERGO®) on skin parameters in healthy adult women aged 35–59 years who reported subjective signs of skin aging. Objective measurements including melanin and erythema indices, skin glossiness, elasticity, and wrinkle and pigmentation counts were used to comprehensively evaluate changes in skin condition.
The OCTN1 transporter is preferentially expressed in basal and granular epidermal layers, where cellular renewal and barrier maintenance are most active. Once internalized, ergothioneine localizes to mitochondria, where it directly scavenges reactive oxygen species (ROS) and protects mitochondrial DNA from UV- and inflammation-induced damage.
At the signaling level, ergothioneine activates key protective pathways such as the Nrf2/ARE axis, enhancing expression of antioxidant enzymes including HO-1, NQO1, and γ-GCLC. These enzymes contribute to redox homeostasis and glutathione regeneration, reinforcing cellular defense systems against photoaging and environmental insult.
In parallel, ergothioneine modulates the PI3K/Akt/Nrf2 and SIRT1/Nrf2 pathways, which are implicated in collagen preservation, inflammation resolution, and mitochondrial maintenance. These pathways converge to reduce matrix metalloproteinase (MMP) activity, enhance collagen synthesis, and suppress pro-inflammatory cytokines (TNF-α, IL-6, IL-1β), all of which are central to maintaining skin structure and function.
Compared to placebo, the DR.ERGO® ergothioneine group showed significantly greater improvements in melanin and erythema reduction, skin glossiness, elasticity, and wrinkle and spot reduction. No adverse events were reported.
These findings corroborate and extend previous clinical evidence from (Hanayama et al., 2024), who investigated an ergothioneine-rich mushroom extract (Pleurotus sp., 25 mg ergothioneine/day) in a 12-week randomized double-blind trial, and (Chunyue Zhang, 2023), who examined pure ergothioneine supplementation (25 mg/day) in a 4-week open-label study. We contextualized our results within this existing literature by comparing key outcomes.
Several limitations should be acknowledged:
The study cohort consisted solely of Japanese women aged 35–59 years, which may limit generalizability across sexes, ethnicities, and age groups.
The 8-week intervention period, while sufficient to detect short-term effects, does not allow conclusions about the sustainability of benefits or the risk of relapse upon discontinuation.
The placebo group also showed modest improvements in self-perception, highlighting the well-documented placebo response in beauty and wellness studies.
This study focused on a single daily dosage (30 mg/day) without evaluating dose–response relationships, and hydration-specific endpoints such as corneometry or transepidermal water loss (TEWL) were not included.”
Two clinical trials investigated ergothioneine’s effects on sleep quality:
“A four-week administration of 20 mg/day ergothioneine (EGT), a strong antioxidant, improves sleep quality; however, its effect at lower doses remains unclear. This study estimated the lower effective doses of EGT using a physiologically based pharmacokinetic (PBPK) model in two clinical trials.
In Study 1, participants received 5 or 10 mg/day of EGT for 8 weeks, and their plasma and blood EGT concentrations were measured. An optimized PBPK model incorporating absorption, distribution, and excretion was assembled. Our results showed that 8 mg/day of EGT for 16 weeks was optimal for attaining an effective plasma EGT concentration.
In Study 2, a randomized, double-blind, placebo-controlled study, participants received 8 mg/day EGT or a placebo for 16 weeks. The subjective sleep quality was significantly improved in the EGT group than in the placebo group.
In mammals, EGT is not generated in the body but is acquired from the diet via the carnitine/organic cation transporter OCTN1/SLC22A4. Its plasma concentration after oral administration is quite stable and gradually increases after repeated dosing on a multi-day basis.
Blood concentrations of EGT increase after Day 8 when EGT intake is interrupted, and they continue to increase until Day 35. The delayed increase in EGT concentration in the blood, compared with that in the plasma, can be interpreted as its efficient uptake by undifferentiated blood cells, which express high levels of OCTN1/SLC22A4 in the bone marrow, and subsequent differentiation to mature blood cells that enter the circulation. This may imply the nonlinear absorption, distribution, and excretion of EGT owing to saturation of the transporter at higher concentrations, potentially leading to difficulty in model construction.
This is the first study to propose a strategy to estimate lower effective doses based on the PBPK model.”
The bolded section above referenced a 2016 study / third graphic for Ergothioneine dosing, Part 2, where a 25 mg dosing stopped after Day 7, but the plasma ergothioneine level stayed high through Day 35. I didn’t see that the referenced 2004 and 2010 studies addressed this 2016 finding.
I also didn’t see that this study’s mathematical model accounted for saturation of the OCTN1 transporter or other effects, such as a very small ergothioneine clearance rate. Okay, lower the ergothioneine dose, and achieve a lower persistent plasma ergothioneine level, to what benefit?
“The present study demonstrated that OCTN1 is associated with myeloid cells rather than lymphoid cells, and especially with erythroid-lineage cells at the transition stage from immature erythroid cells to peripheral mature erythrocytes.”
Persistent high ergothioneine levels aren’t costless. Skewing bone marrow stem cells and progenitor cells toward a myeloid lineage is done at the expense of lymphocytes, T cells, B cells, and other lymphoid lineages.
Where are the studies that examine these tradeoffs? Subjective sleep quality in this study and sleep initiation in the first study above aren’t sufficiently explanatory.
A study investigated associations of plasma ergothioneine levels and cognitive changes in older adults over a two-year period:
“Observational studies have found that lower plasma levels of ergothioneine (ET) are significantly associated with higher risks of neurodegenerative diseases. However, several knowledge gaps remain:
Most of the above studies were based on cross-sectional study design, and potential reverse causation cannot be excluded. It has been suggested that plasma ET declines concomitantly with the deterioration of cognitive function.
Since the impact of a single dietary factor on health is mild, it is prone to be affected by the baseline characteristics of subjects (such as sex, educational level, disease status and gene polymorphism). However, no study has systematically evaluated potential effect modifiers on the association between ET levels and cognitive function.
The dose-response distribution between ET and cognitive function remains undetermined.
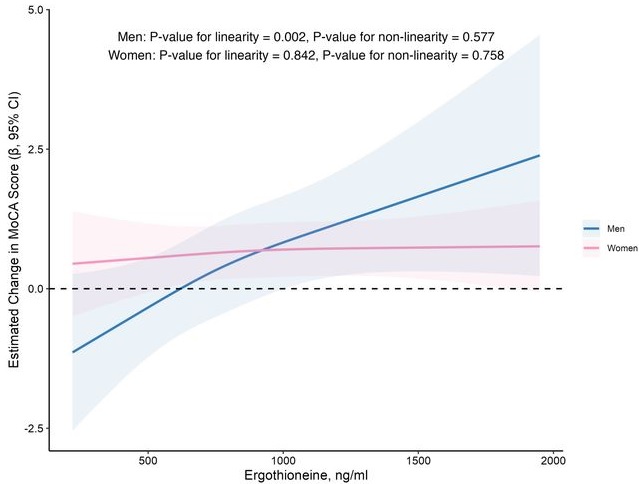
In this prospective cohort study of 1,131 community-dwelling older adults (mean age 69 years), higher baseline plasma ET levels were significantly associated with slower cognitive decline, as assessed by Montreal Cognitive Assessment (MoCA) scores, during a 2-year follow-up period.
When the plasma concentration of ET exceeds 1,000 ng/mL, the decline in cognitive function significantly slows down. However, this association has only been observed in men.
Domain-specific analysis found that the observed ET-MoCA association was mainly driven by the temporary slowdown in the decline of visuospatial/executive and delayed recall. Impaired delayed recall represents one of the earliest and most sensitive cognitive markers of dementia progression, predictive of conversion from MCI to dementia. The preferential preservation of this function by ET suggests targeted neuroprotective effects within the hippocampus.
Visual inspection of the spline curves revealed a potential plateauing effect at ET concentrations ≥1,000 ng/mL in the total population.
Baseline ET concentrations differed between men and women. Most men (81.5%) had concentrations below 1,000 ng/mL (median 754.2, IQR 592.0–937.9 ng/mL). Women exhibited substantially higher median plasma ET concentrations than men, with 35.7% of women exceeded 1,000 ng/mL (median 890.1, IQR 709.7–1,095.6 ng/mL).
Our study included only participants with normal cognitive function, and the results remained robust even after excluding those with baseline cognitive function at the lower end of the normal range. Collectively, our findings support that low ET intake occurs prior to cognitive decline.
Our findings indicate that higher plasma ET levels are significantly associated with slower cognitive decline independent of confounders in non-demented community-dwelling elderly participants, with such association observed in men but not women. Dose-response curves indicated plateauing effects above 1000 ng/mL.”
The average age of this study and the first trial above were both 69 years. Since the first trial’s participants showed slightly above-average cognitive function (neurocognitive index median = 105), with plasma ergothioneine levels of median = 1154 nM at baseline, and this study showed plateauing effects above 1000 ng/mL, I wonder how raising plasma ergothioneine levels above 1000 ng/mL could possibly show a net benefit for older people? What are the trade-offs for older people between potentially increasing slightly above-average cognitive function with ergothioneine and its other effects from saturating their OCTN1 transporter?
This study is at its preprint stage. I’m interested to see if its peer review prompts these researchers to also investigate the common finding that people who are most deficient at baseline have the greatest improvements. If so, would these sex-specific associations still hold?
Wrapping up with a study that investigated associations of serum ergothioneine levels with the risk of developing dementia:
“1344 Japanese community-residents aged 65 years and over, comprising 765 women and 579 men, without dementia at baseline were followed prospectively for a median of 11.2 years.
During follow-up, 273 participants developed all-cause dementia. Among them, 201 had Alzheimer’s disease (AD) and 72 had non-Alzheimer’s disease (non-AD) dementia.
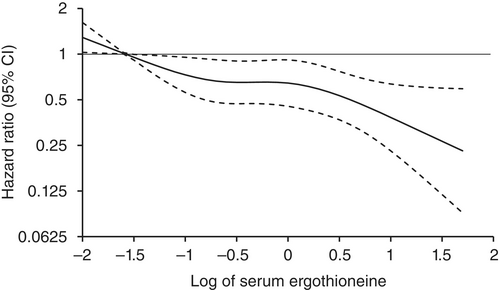
Age- and sex-adjusted hazard ratios (HRs) for all-cause dementia, AD, and non-AD dementia decreased progressively across increasing quartiles of serum ergothioneine. These associations remained significant after adjustment for a wide range of cardiovascular, lifestyle, and dietary factors, including daily vegetable intake.
In subgroup analysis, association between serum ergothioneine levels and the risk of dementia tended to be weaker in older participants and in women:
In older individuals, cumulative burden of multiple risk factors such as hypertension, diabetes mellitus, and smoking may contribute to both neurodegenerative and vascular pathology, potentially diminishing the relative influence of ergothioneine.
In women, postmenopausal hormonal changes, particularly the decline in estrogen, have been associated with increased oxidative stress and a higher vulnerability to neurodegenerative changes.
Several limitations should be noted:
Since serum ergothioneine levels and other risk factors were measured only at baseline, we could not evaluate the changes of serum ergothioneine levels during the follow-up period. Lifestyle modifications during follow-up could have influenced serum ergothioneine levels and other risk factors. In addition, serum ergothioneine level was measured only once, and from a sample.
We cannot rule out residual confounding factors, such as other nutrients in mushrooms and socioeconomic status.
There is a possibility that dementia cases at the prodromal stage were included among participants with low serum ergothioneine levels at baseline.
We are unable to specify which mushroom varieties were predominantly consumed by participants in the town of Hisayama.
Given the limited discriminative ability of serum ergothioneine and potential degradation due to long-term sample storage, we were unable to explore a clinically meaningful threshold value of serum ergothioneine.
Generalizability of findings was limited because participants of this study were recruited from one town in Japan.
These findings suggest that the potential benefit of ergothioneine may be attenuated in individuals with pre-existing, multifactorial risk profiles for dementia.
Our findings showed that higher serum ergothioneine levels were associated with a lower risk of developing all-cause dementia, AD, and non-AD dementia in an older Japanese population. Since ergothioneine cannot be synthesized in the human body, a diet rich in ergothioneine may be beneficial in reducing the risk of dementia.”
For five years I got most of my estimated 7 mg daily ergothioneine intake from mushrooms in AGE-less chicken vegetable soup per Ergothioneine dosing. The soup was always boring, but I got too bored this year and stopped making it. I haven’t replaced mushroom intake with supplements.
I still don’t eat fried or baked foods, preferring sous vide and braising cooking methods to avoid exogenous advanced glycation end products. I avoid buying foods that evoke a hyperglycemic response or otherwise form excessive endogenous AGEs per All about AGEs.
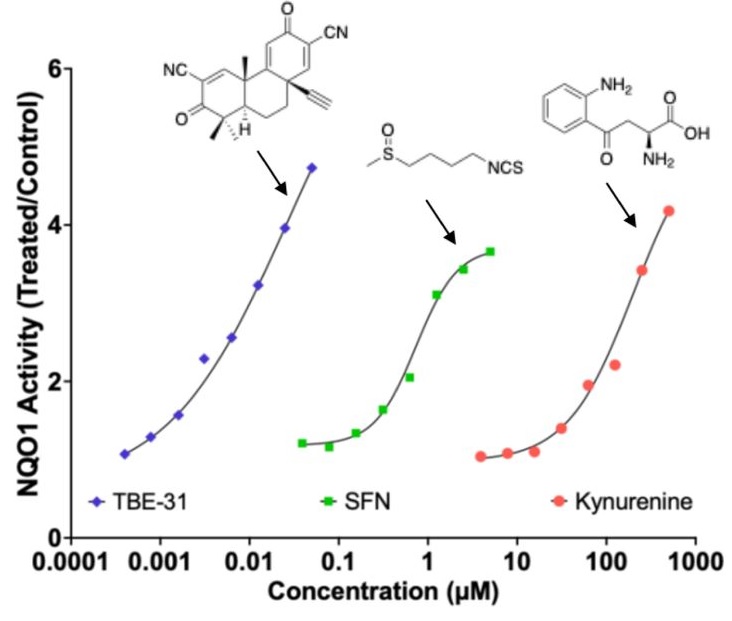
Here are two 2025 papers, starting with a rodent study that investigated interactions between the Nrf2 and kynurenine pathways:
“Exposure to the tryptophan metabolite kynurenine and its electrophilic derivative kynurenine-carboxyketoalkene (Kyn-CKA) leads to an increase in the abundance of transcription factor Nrf2 and induction of Nrf2-target genes. The Keap1/Nrf2 system is the main orchestrator of cellular defence against environmental stress, most notably oxidative and inflammatory stress.
Nrf2 can be activated pharmacologically by small molecules, the majority of which are electrophiles and oxidants that modify specific cysteine-based sensors in Keap1. C151 in Keap1 is the target of the isothiocyanate sulforaphane, a classical Nrf2 activator that has been employed in ∼90 clinical trials, as well as for the two Nrf2 activators that are clinically in use: dimethyl fumarate, for relapsing remitting multiple sclerosis, and omaveloxolone, for Friedreich’s ataxia.
Kynurenine is an endogenous metabolite derived from the essential amino acid tryptophan. Kynurenine and its metabolites, such as the electrophilic kynurenine-carboxyketoalkene (Kyn-CKA), have been demonstrated to activate Nrf2 in other pathologies, including sickle cell disease, attenuating inflammation. Moreover, identification of the gene encoding the kynurenine-metabolising enzyme kynureninase as a gene transcriptionally upregulated by Nrf2, provides a plausible negative feedback regulatory mechanism.
Because kynurenine is not electrophilic, whereas its metabolite Kyn-CKA is, we considered the possibility that Kyn-CKA is the actual Nrf2 activator. Using biochemical and cell-based assays, we found that Kyn-CKA reacts with C151 in the BTB domain of Keap1 and increases the thermostability of Keap1, indicating target engagement. Consequently, Nrf2 accumulates and induces transcription of antioxidant/electrophile-responsive element (ARE/EpRE)-driven genes.
These findings demonstrate that Kyn-CKA targets C151 in Keap1 to derepress Nrf2, and reveal that Nrf2 is a main contributor to the anti-inflammatory activity of Kyn-CKA in macrophages.”
A review subject was targeting nicotinamide adenine dinucleotide, oxidized form (NAD+) for clinical use:
“Mammalian NAD+ biosynthesis includes four known pathways, primarily occurring in cytoplasm:
(a) the NRH pathway;
(b) the salvage pathway;
(c) the Preiss–Handler pathway; and
(d) the kynurenine pathway.
The de novo kynurenine pathway metabolizes tryptophan (Trp) to NAD+, producing various intermediates that serve as biomarkers for different diseases. These intermediates show alterations in various pathological conditions.
While kynurenine and its metabolic derivatives are intermediates in the de novo NAD+ biosynthesis pathway, these are also produced independently in various physiological contexts, particularly in immune cells, where they act as immunomodulatory compounds.”
This second paper above showed a graphic of the Nrf2 and kynurenine pathways together in a diagram showing relationships between NAD+ augmentation and the hallmarks of aging, but didn’t elaborate other than labeling their box Dysbiosis. So how these two pathways interact is better outlined in the first paper above with explaining how a kynurenine-metabolizing enzyme is one of the hundreds of Nrf2 target genes, creating a natural feedback loop between Nrf2 activation and the kynurenine pathway.
These reviewers also lumped SIRT1 in their Dysbiosis box, and into several other boxes, probably due to the penultimate coauthor’s influence:
However, repeating something over and over doesn’t make it scientifically valid regardless of the number of citations. Or, as a 2022 review Sirtuins are not conserved longevity genes concluded:
“A global pursuit of longevity phenotypes was driven by a mixture of framing bias, confirmation bias, and hype. Review articles that propagate these biases are so rampant that few investigators have considered how weak the case ever was for sirtuins as longevity genes.
Acknowledging that a few positive associations between sirtuins and longevity have been identified after thousands of person-years and billions of dollars of effort, we review the data and suggest rejection of the notions that sirtuins (i) have any specific connection to lifespan in animals and (ii) are primary mediators of the beneficial effects of NAD repletion.”
Continuing Part 1 with three 2025 papers, starting with a rodent study of dietary mussel plasmalogens’ effects on atherosclerosis:
“The purpose of this study was to clarify the underlying mechanisms of Mytilus edulis-derived plasmalogens (Pls) against atherosclerosis (AS) in ApoE−/− mice induced by a high-fat diet (HFD), through a comprehensive analysis of hepatic metabolomics and aortic transcriptomics data. Besides Pls role as the storage pool of n-3 PUFAs, the structural feature of vinyl ether bond at sn-1 position confers multiple advantages upon Pls compared to their diacyl counterparts, including enhanced antioxidant capacity, increased membrane fluidity, as well as improved stability and stability of biomembranes.
The C57BL/6 mouse strain is susceptible to high-fat diet (HFD)-induced AS lesions, and ApoE knockout accelerates AS development. Molecular mechanisms by which Pls ameliorate AS were investigated through a comprehensive analysis of hepatic metabolomics and aortic transcriptome profiles, focusing on changes in gene related to the p38 mitogen-activated protein kinase (MAPK) signaling pathway and the downstream inflammatory response.
The concentration of Pls in mussel tissues is 32 μgmg−1 (dry weight), and the obtained Pls contains 49.53% of phosphatidylethanolamine-Pls, 35.87% of phosphatidylcholine-Pls, and 14.60% of phosphatidylserine-Pls. The main fatty acid compositions of Pls are presented in Supplementary Table 1, which indicates that EPA accounts for 45.82% and the n-3/n-6 ratio is 3.84.
Pls inhibited aortic lipid accumulation, prevented thickening of the aortic wall, and suppressed collagen accumulation at the aortic-heart junction. Pls inhibited HFD-induced loosening of hepatocyte arrangement, vacuolization, and accumulation of lipid droplets.
Although several key components of MAPK signaling pathway were suppressed at both the transcriptional and protein levels in Pls-treated mice, no significant changes in phosphorylated p38 protein were observed among the experimental groups in our study. Further research is needed to elucidate the overall inhibitory mechanism of Pls on p38 protein and the MAPK signaling pathway.”
A rodent / human cell study investigated effects of plasmalogens in innate immune system macrophages on atherosclerosis:
“We demonstrate that simultaneous inactivation of two key enzymes involved in macrophage polyunsaturated fatty acid (PUFA) metabolism—ELOVL5, which elongates long-chain PUFAs, and LPCAT3, which incorporates them into phospholipids—disrupts membrane organization by promoting the formation of cholesterol-enriched domains. This increases macrophage sensitivity to cytotoxic oxysterols and leads to more vulnerable atherosclerotic plaques with enlarged necrotic cores in a mouse model of atherosclerosis.
We identified ELOVL5 as one elongase facilitating the conversion of C20 to C22 PUFA. In humans, analysis of 187 carotid plaques reveals a positive correlation between LPCAT3/ELOVL5-generated phospholipids—including arachidonate (C20:4 n-6)-containing ether lipids—and more stable plaque profiles. Additionally, Mendelian randomization analysis supports a causal relationship between LPCAT3 expression and reduced risk of ischemic stroke.
Potentially beneficial effects we observed in mice and in human atheroma plaques were mainly associated with PLs enriched in omega-6, particularly in AA. Although omega-6 FAs are often considered as pro-inflammatory, their role is undergoing reconsideration, with markers linked to the intake of omega-6 appearing beneficial in the context of cardiovascular diseases. In this context, it is worth to note that AA-containing plasmalogens have been previously identified as markers of healthy obesity.
Our findings uncover a regulatory circuit essential for PUFA-containing phospholipid generation in macrophages, positioning PUFA-containing ether lipids as promising biomarkers and therapeutic targets.”
A human study included plasmalogens in investigating associations among people with mental illness and their lipid profiles:
“Plasma lipidomic profiles of 623 individuals (188 schizophrenia (SCZ), 243 bipolar disorder (BD), 192 healthy controls) belonging to the PsyCourse Study were assessed using liquid chromatography and untargeted mass spectrometry. Exact etiology of these major mental health disorders is yet unknown and while their symptoms overlap, their diagnostic criteria are based on clinical evaluations of symptoms without objective markers.
Cognitive dysfunction is among the most disabling symptoms of SCZ and BD, and is difficult to treat with the commonly used pharmacologic regimes. Consequently, it has important impacts on long-term functional outcomes.
We aimed to answer the question, whether specific lipid species or classes were associated with differential performance across various cognitive domains, including psychomotor and processing speed, executive function, short-term and working memory and crystalized intelligence and whether these associations were affected by diagnoses.
Lipids belonging to the phosphatidylethanolamine plasmalogen (PE-P) class emerged as the main lipid class associated negatively with DG-SYM test performance, representative of processing and psychomotor speed. Our findings showed that higher levels of PE-P 42:5, PE-P 40:4, PE-P 40:5, and ceramide 38:1 in plasma samples of our study are significantly associated with poorer DG-SYM test performance. The DG-SYM test mainly measures processing speed, the amount of time required to complete a series of cognitive tasks. Enrichment analysis also showed significant associations between other lipid classes and various cognitive tests.
Our findings suggest a link between lipids and cognitive performance independent of mental health disorders. Independent replication is warranted to better understand if phosphatidylethanolamines could represent an actionable pharmacologic target to tackle cognitive dysfunction, an important unmet clinical need that affects long-term functional outcomes in individuals with severe mental health disorders.”
It was apparently beyond these researchers’ expertise to offer informed discussion on this study’s associative link between enrichment of these three phosphatidyl ethanolamine plasmalogens and cognitive dysfunction. Grok countered that their depletion was associated with neurodegenerative diseases (Alzheimer’s, Parkinson’s, multiple sclerosis), cardiovascular risk / oxidized-LDL burden, and chronic fatigue / post-viral syndromes.
A 2025 review subject was sulforaphane and brain health. This paper was the latest in a sequence where the retired lead author self-aggrandized his career by citing previous research.
He apparently doesn’t personally do what these research findings suggest people do. The lead author is a few weeks older than I am, and has completely white hair per an interview (Week 34 comments). I’ve had dark hair growing in (last week a barber said my dark hair was 90%) since Week 8 of eating broccoli sprouts every day, which is a side effect of ameliorating system-wide inflammation and oxidative stress.
If the lead author followed up with what his research investigated, he’d have dark hair, too. Unpigmented white hair and colored hair are both results of epigenetics.
Contrast this lack of personal follow-through of research findings with Dr. Goodenowe’s protocol where he compared extremely detailed personal brain measurements at 17 months and again at 31 months. He believes enough in his research findings to personally act on them, and demonstrate to others how personal agency can enhance a person’s life.
It’s every human’s choice whether or not we take responsibility for our own one precious life. I’ve read and curated on this blog many of this paper’s references. Five years ago for example:
Two 2024 reviews by the same group that published Sulforaphane in the Goldilocks zone investigated dietary polyphenols’ effects as “hormetic nutrients”:
“Polyphenols display biphasic dose–response effects by activating at a low dose the Nrf2 pathway resulting in the upregulation of antioxidant vitagenes [see diagram]. We aimed to discuss hormetic nutrients, including polyphenols and/or probiotics, targeting the Nrf2 pathway and vitagenes for the development of promising neuroprotective and therapeutic strategies to suppress oxidative stress, inflammation and microbiota deregulation, and consequently improve cognitive performance and brain health.
Hormetic nutrition through polyphenols and/or probiotics targeting the antioxidant Nrf2 pathway and stress resilient vitagenes to inhibit oxidative stress and inflammatory pathways, as well as ferroptosis, could represent an effective therapy to manipulate alterations in the gut microbiome leading to brain dysfunction in order to prevent or slow the onset of major cognitive disorders. Notably, hormetic nutrients can stimulate the vagus nerve as a means of directly modulating microbiota-brain interactions for therapeutic purposes to mitigate or reverse the pathophysiological process, restoring gut and brain homeostasis, as reported by extensive preclinical and clinical studies.”
I’m not onboard with this study’s probiotic assertions because most of the cited studies contained unacknowledged measurement errors. Measuring gut microbiota, Part 2 found:
“The fecal microbiome does not represent the overall composition of the gut microbiome. Despite significant roles of gut microbiome in various phenotypes and diseases of its host, causative microbes for such characteristics identified by one research fail to be reproduced in others.
Since fecal microbiome is a result of the gut microbiome rather than the representative microbiome of the GI tract of the host, there is a limitation in identifying causative intestinal microbes related to these phenotypes and diseases by studying fecal microbiome.”
These researchers also erroneously equated isothiocyanate sulforaphane’s Nrf2-activating mechanisms with polyphenols activating Nrf2.
This research group did better in clarifying polyphenols’ mechanisms in a review of hormetic dose-response effects of the polyphenol rosmarinic acid:
“This article evaluates whether rosmarinic acid may act as a hormetic agent, mediating its chemoprotective effects as has been shown for similar agents, such as caffeic acid, a derivative of rosmarinic acid.
Rosmarinic acid enhanced memory in institute of cancer research male mice in the Morris water maze (escape latency).
Of importance in the evaluation of rosmarinic acid are its bioavailability, metabolism, and tissue distribution (including the capacity to affect and/or cross the BBB and its distribution and half-life within the brain). In the case of polyphenols, including rosmarinic acid, they are typically delivered at low doses in the diet and, in most instances, they do not escape first-pass metabolism, with the prominent chemical forms being conjugates of glucuronides and sulfates, with or without methylation.
These conjugated metabolites are chemically distinct from the parent compound, showing considerable differences in size, polarity, and ionic form. Their biological actions are quite different from the parent compound.
Bioavailability studies reveal that maximum concentrations in plasma typically do not exceed 1 µM following consumption of 10–100 mg of a single phenolic compound, with the maximum concentration occurring typically less than 2 h after ingestion, then dropping quickly thereafter. In the case of the in vitro studies assessed herein, and with few exceptions, most of the studies employed concentrations >10 µM with some studies involving concentrations in the several hundred µM range, with the duration of exposure typically in the range of 24–72 h, far longer duration than the very short time interval of a few minutes to several hours in human in vivo situations.
We strongly recommend that all experiments using in vitro models to study biological responses to dietary polyphenols use only physiologically relevant flavonoids and their conjugates at appropriate concentrations, provide evidence to support their use, and justify any conclusions generated. When authors fail to do this, referees and editors must act to ensure that data obtained in vitro are relevant to what might occur in vivo.”
A 2024 primate study investigated effects of an off-patent drug on age-related changes:
“We evaluated geroprotective effects of metformin on adult male cynomolgus monkeys. The study encompassed a comprehensive suite of physiological, imaging, histological, and molecular evaluations, substantiating metformin’s influence on delaying age-related phenotypes at the organismal level.
Results highlighted a significant slowing of aging indicators, notably a roughly 6-year regression in brain aging. Metformin exerts a substantial neuroprotective effect, preserving brain structure and enhancing cognitive ability.
Geroprotective effects on primate neurons were partially mediated by activation of Nrf2, a transcription factor with anti-oxidative capabilities.”
“Metformin treatment resulted in increased nuclear phosphorylated Nrf2, accompanied by up-regulation of Nrf2 target genes like HO-1, NQO-1, SOD3, GPX2, and GPX1, which were generally suppressed and typically down-regulated during human neuron senescence.
Genes pivotal for neuronal function, such as dendrite morphogenesis/extension and synapse assembly (e.g., GSK3B, GRID2, and NRG3), were down-regulated during aging in excitatory neurons (ExN), inhibitory neurons (InN), oligodendrocytes (OL), oligodendrocyte progenitor cells (OPC), microglia, and astrocyte but were restored by metformin treatment. By contrast, pathways that were up-regulated during aging, including activation of the immune response, complement activation, and regulation of the TGF-b receptor signaling pathway, were reset to lower levels by metformin treatment.
We verified that markers associated with brain aging and progression of neurodegenerative diseases were restored by metformin treatment to levels similar to those observed in young monkeys. Additionally, we observed that reduced myelin sheath thickness, a characteristic of aged monkeys, was rebuilt to a younger state following metformin treatment.
These findings align with the levels of nuclear-localized phosphorylated Nrf2, suggesting that Nrf2 pathway activation is a key mechanism in metformin’s role in delaying human neuronal aging and, by extension, brain aging. Consistent with our in vitro findings, Nrf2 pathway activation was also detected across multiple tissues in metformin-treated monkeys, including frontal lobe neurons.
At last count, I’ve curated 250+ papers this decade on cruciferous vegetables, and many of these explored relationships with Nrf2 activation. Basically, eating a clinically-relevant daily dose of 3-day-old cruciferous sprouts and taking off-patent metformin both induce Nrf2 activation effects.
Don’t expect to see many researchers highlighting this equivalency. They’d rather wait another decade to nitpick other studies with not-enough-subjects / not-exactly replicated / other nitpicks before expressing opinions urging caution from their nursing home beds.
But even then, they won’t get their facts straight. For example, a contemporaneous opinion article https://www.nature.com/articles/d41586-024-02938-w “The brain aged more slowly in monkeys given a cheap diabetes drug” attempted to summarize this study, and flubbed two points:
1. The study said: “We conducted a proof-of-concept study involving male cynomolgus monkeys (Macaca fascicularis) aged between 13 and 16 years, roughly equivalent to approximately 40–50 years in humans. Monkeys adhered to this regimen for a period of 1,200 days, approximately 3.3 years, which corresponds to about 10 years in humans.”
The opinion claimed: “Animals took the drug for 40 months, which is equivalent to about 13 years for humans.”
2. The opinion quoted a New York City researcher involved in a separate metformin study and employed at a medical school for:
“Research into metformin and other anti-ageing candidates could one day mean that doctors will be able to focus more on keeping people healthy for as long as possible rather than on treating diseases.”
This statement is a big break from the realities of medical personnel daily actions at least so far this decade, which is when I started to pay close attention:
Doctors have very little diet and exercise training in medical school. There’s no way they can give health advice. There’s no way that a “keeping people healthy”paradigm will emerge from the current medical system.
Fixing a disease doesn’t restore a patient’s health. Dr. (PhD) Goodenowe cites several examples in his talks, such as a study that compared colorectal cancer therapy with post-operation patient health.
If you listen to yesterday’s two-hour-long podcast, the currently injured person in the first hour gave plenty of contrary evidence of doctors’ focuses: behaviors of trying to blame and gaslight the patient, thinly-disguised punitive actions, CYA etc., all of which they will be sued for one day. The doctor in the second hour provided an example of the quoted researcher in her explanation of how doctors higher in the hierarchy either can’t see or can’t admit realities of doctor/patient interactions, and what therapies have actually benefited or harmed a patient.
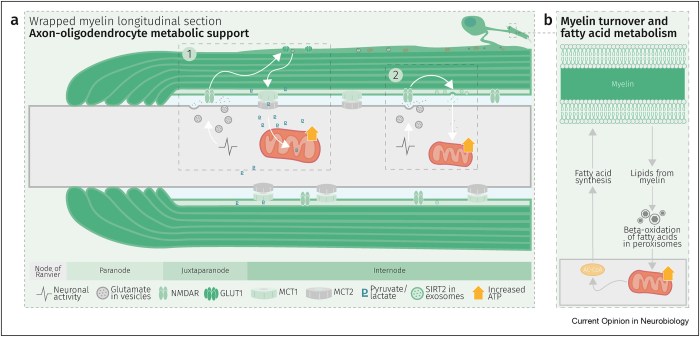
Three papers on myelin and oligodendrocytes, starting with a 2023 review:
“Myelin is the spiral ensheathment of axons by a lipid and cholesterol-rich glial cell membrane that reduces capacitance and increases resistance of the axonal membrane. Axonal myelination speeds up nerve conduction velocity as a function of axon diameter.
While myelination proceeds rapidly after birth in the peripheral nervous system, central myelination is a spatially and temporally more regulated process. Ongoing myelination of the human brain has been documented at up to 40 years of age. This late myelination in the adult cortex is followed by exhaustion of oligodendrocyte precursor cells (OPC) with senescence and a gradual loss of myelin integrity in the aging brain.
The brain is well known for its high energy demands, specifically in gray matter areas. In white matter tracts, energy consumption is lower. Myelination poses a unique challenge for axonal energy generation where myelin sheaths cover more than 95% of the axonal surface areas.
Oligodendrocytes help support axonal integrity. Oligodendrocytes survive well in the absence of mitochondrial oxidative phosphorylation, and without signs of myelin loss, cell death, neurodegeneration or secondary inflammation.
Glycolysis products of oligodendroglial origin are readily metabolized in axonal mitochondria. Oligodendroglial metabolic support is critical for larger and faster-spiking myelinated axons that also have a higher density of mitochondria. An essential requirement for the direct transfer of energy-rich metabolites from oligodendrocytes to the myelinated axonal compartment is ‘myelinic channels’ within the myelin sheath.
Interactions of oligodendrocytes and myelin with the underlying axon are complex and exceed the transfer of energy-rich metabolites. Continuous turnover of myelin membranes by lipid degradation and fatty acid beta-oxidation in mitochondria and peroxisomes leads to recycling of acetate residues by fatty acid synthesis and membrane biogenesis.
In human multiple sclerosis (MS) and its animal model myelin oligodendrocyte glycoprotein-experimental autoimmune encephalomyelitis (MOG-EAE), acute inflammatory demyelination is followed by axonal degeneration in lesion sites that is mechanistically not fully understood. It is widely thought that demyelination and the lack of an axon-protective myelin sheath in the presence of numerous inflammatory mediators are the main causes of axon loss.
But unprotected axons improve rather than worsen the overall clinical phenotype of EAE mice which exhibited the same degree of autoimmunity. Thus, ‘bad myelin is worse than no myelin’ because MS-relevant myelin injuries perturb the integrity of myelinic channels and metabolic support.
Dysfunctional or injured oligodendrocytes that do not allow for compensation by any other cell types turn the affected myelin ensheathment into a burden of the underlying axonal energy metabolism, which causes irreversible axon loss. Any loss of myelin integrity, as seen acutely in demyelinating disorders or more gradually in the aging brain, becomes a risk factor for irreversible neurodegeneration.”
A 2024 review focused on myelin and oligodendrocyte plasticity:
“This review summarizes our current understanding of how myelin is generated, how its function is dynamically regulated, and how oligodendrocytes support the long-term integrity of myelinated axons.
Apart from its unique ultrastructure, there are several other exceptional features of myelin. One is certainly its molecular composition. Another is its extraordinary stability. This was compellingly illustrated when 5000-year-old myelin with almost intact ultrastructure was dissected from a Tyrolean Ice Man.
Myelin is a stable system in contrast to most membranes. However, myelin is compartmentalized into structurally and biochemically distinct domains. Noncompacted regions are much more dynamic and metabolically active than tightly compacted regions that lack direct access to the membrane trafficking machinery of oligodendrocytes.
The underlying molecular basis for stability of myelin is likely its lipid composition with high levels of saturated, long chain fatty acids, together with an enrichment of glycosphingolipids (∼20% molar percentage of total lipids) and cholesterol (∼40% of molar percentage of total lipids). In addition, myelin comprises a high proportion of plasmalogens (ether lipids) with saturated long-chain fatty acids. In fact, ∼20% of the fatty acids in myelin have hydrocarbon chains longer than 18 carbon atoms (∼1% in the gray matter) and only ∼6% of the fatty acids are polyunsaturated (∼20% in gray matter).
With maturation of oligodendrocytes, the plasma membrane undergoes major transformations of its structure. Whereas OPCs are covered by a dense layer of large and negatively charged self-repulsive oligosaccharides, compacted myelin of fully matured oligodendrocytes lacks most of these glycoprotein and complex glycolipids.
Schematic depiction of an oligodendrocyte that takes up blood-derived glucose and delivers glycolysis products (pyruvate/lactate) via monocarboxylate transporters (MCT1 and MCT2) to myelinated axons. Oligodendrocytes and myelin membranes are also coupled by gap junctions to astrocytes, and thus indirectly to the blood–brain barrier.
Adaptive myelination refers to dynamic events in oligodendroglia driven by extrinsic factors such as experience or neuronal activity, which subsequently induces changes in circuit structure and function. Understanding how these adaptive changes in neuron-oligodendroglia interactions impact brain function remains a pressing question for the field.
Transient social isolation during adulthood results in chromatin and myelin changes, but does not induce consequent behavioral alterations. When mice undergo a social isolation paradigm during early life development, they similarly exhibit deficits in prefrontal cortex function and myelination, but these deficiencies do not recover with social reintroduction. This implicates a critical period for social deprivation effects on myelin dynamics. Experience-dependent changes in myelin dynamics may depend on not only the age, brain region, and cell type studied, but also the specific myelin structural change assessed.
Local synaptic neurotransmitter release along an axon not only affects the number of OPCs and oligodendrocytes associated with that axon and local synthesis of myelin proteins, but also drives preferential selection of active axons for myelination over the ensheathment of electrically silenced neighboring axons. Neuronal activity–induced plasticity may preferentially impact brain regions that remain incompletely myelinated compared to more fully myelinated tracts.
Whereas the myelin sheath has been regarded for a long time as an inert insulating structure, it has now become clear that myelin is metabolically active with cytoplasmic-rich pathways, myelinic channels, for movement of macromolecules into the periaxonal space. The myelin sheath and its subjacent axon need to be regarded as one functional unit, which are not only morphological but also metabolically coupled.”
A 2024 rodent study investigated oligodendrocyte precursor cell transcriptional and epigenetic changes:
“We used single-cell RNA sequencing (scRNA-seq), single-cell ATAC sequencing (scATAC-seq), and single-cell spatial transcriptomics to characterize murine cortical OPCs throughout postnatal life. One group (active, or actOPCs) is metabolically active and enriched in white matter. The second (homeostatic, or hOPCs) is less active, enriched in gray matter, and predicted to derive from actOPCs. Relative to developing OPCs, both actOPCs and hOPCs are less active metabolically and have less open chromatin.
In adulthood, these two groups are transcriptionally but not epigenetically distinct, indicating that they may represent different states of the same OPC population. If that is the case, then one model is that the parenchymal environment maintains adult OPCs within an hOPC state, whereas those OPCs recruited into white matter or exposed to demyelinated axons may transition toward an actOPC state in preparation for making new oligodendrocytes. We do not yet know the functional ramifications of these differences, but this finding has clear implications for the development of therapeutic strategies for adult remyelination.
Another finding is that developing but not adult actOPC chromatin is preferentially open for binding motifs associated with neural stem cells, transit-amplifying precursors, and neurogenesis. Although this may simply reflect their origin as the immediate progeny of neonatal neural precursor cells, it may also explain why developing but not adult OPCs have the capacity to make neurons in culture.
If we could, at least in part, reverse the global chromatin shutdown that occurs between development and adulthood, then perhaps adult OPCs may reacquire the ability to make neurons or become better able to generate new oligodendrocytes for remyelination.”
A free educational series recorded in 2021-2022 available at https://drgoodenowe.com/dr-goodenowes-educational-seminars/ takes the viewer through underlying research and principles of Dr. Goodenowe’s approach to health. It’s advertised as lasting four hours, but took me two days to view.
The series’ discussions and references are background material to better understand later presentations and interviews. Points of interest included:
Seminar B100 shows that the metabolomic profile of people who regularly eat broccoli is different than others.
B109 clarifies how peroxisomal function is improved through resistance exercise and intermittent fasting.
C103 and C104 show how plasmalogens act against neurodegeneration (Parkinson’s disease and multiple sclerosis).
Texts below videos are additional information, not transcripts. C101 text is historically informative.
The B200 ProdromeScan tutorial will take more study. But unlike Labcorp tests, ordering a ProdromeScan requires using a practitioner in Dr. Goodenowe’s network.
I sent the following to Prodrome customer service earlier this month:
Please add me to your approved list for ProdromeScan.
Customer service replied:
“We only add health professionals to an approved list, not individuals.”
I responded:
Good morning. I looked at the websites of doctors who are associated with Dr. Goodenowe who are near me. All of them are too compromised for me to establish a doctor / patient relationship. But I’m glad they left up their blog posts from earlier this decade so I could see who they really were before I reached out to them.
I request an exception to the policy.
Customer service replied:
“There is no exception that can be made to this policy. You need to be a patient of a certified practitioner.”
I’ll escalate my request before my 90-day trial of Prodrome Glia and Neuro products ends so I can get an appropriate metabolomic status. Right now, I won’t involve someone I can’t trust just to know my ProdromeScan information that’s additional to next week’s Labcorp tests.
My treatment-result metabolomic data is probably not mature today on Day 29 of ProdromeGlia and ProdromeNeuro supplementation, resistance exercise, and intermittent fasting. I otherwise wouldn’t have experienced these two events:
I’m happy that after starting normal ProdromeGlia and ProdromeNeuro doses at Day 1, then taking a ProdromeGlia loading dose starting from Day 11, my sense of smell and taste improved by Days 13-14.
“We use NAC in the lab all the time because it stops an Nrf2 activation. So that weak pro-oxidant signal that activates Nrf2, you switch it off by giving a dose of NAC. It’s a potent antioxidant in that right, but it’s blocking signalling. And that’s what I don’t like about its broad use.”
If someone bombs themself everyday with antioxidants, they’re doing nothing to improve training of their endogenous systems’ defensive functions. What happens when they stop bombing? One example was a 2022 human study that found GlyNAC-induced improvements dissolved back to baseline after supplements stopped.
“TFEB activity is required for sulforaphane (SFN)-induced protection against both acute oxidant bursts and chronic oxidative stress. SFN-induced TFEB nuclear accumulation was completely blocked by pretreatment of cells by N-acetyl-cysteine (NAC), or by other commonly used antioxidants. NAC also blocked SFN-induced mRNA expression of TFEB target genes, as well as SFN-induced autophagosome formation.”
In this 2023 presentation for a professional audience, Dr. Dayan Goodenowe showed an example of what could be done (in the form of what he personally did at ages 53-54) to restore and augment brain structure and function over a 17-month period by taking plasmalogens and supporting supplements:
Follow the video along with its interactive transcript. Restorative / augmentative supplements included:
Forms of MRI used to document brain structure and function changes were:
Brain volume decreases are the rule for humans beginning at age 40. Dr. Goodenowe documented brain volume increases, which aren’t supposed to happen, but did per the below slide of overall results:
“From a global cortical volume and thickness perspective, 17 months of high-dose plasmalogens reversed ~15 years of predicted brain deterioration.”
Specific increased adaptations in brain measurements over 17 months included:
Cortical thickness .07/2.51 = +3%.
White matter microstructure fractional anisotropy +8%.
Neurovascular coupling signal controlled by noradrenaline integrity.
Amygdala volume +4% and its connectivity to the insula, indicating ongoing anxiety and emotional stress response.
Parahippocampus volume +7%.
Hippocampus fractional anisotropy +5%.
No changes:
Amygdala connectivity to the ventral lateral prefrontal cortex, the same part of the brain that relates to placebo effect.
Hippocampus connectivity.
Decreased adaptations in brain measurements included:
White matter microstructure radial diffusivity -10%.
Amygdala connectivity to the anterior cingulate cortex to suppress / ignore / deny anxiety response.
Amygdala connectivity to the dorsal lateral prefrontal cortex.
Entorhinal cortex volume -14%.
Hippocampus volume -6%.
Hippocampus mean diffusivity (white matter improved, with more and tighter myelin) -4%.
The other half of this video was a lively and wide-ranging Q&A session.
The referenced 2023 study of 653 adults followed over ten years showed what brain deterioration could be expected with no interventions. Consider these annual volume decrease rates to be a sample of a control group:
The daily plasmalogen precursor doses Dr. Goodenowe took were equivalent to 100 mg softgel/kg, double the maximum dose of 50 mg softgel/kg provided during the 2022 clinical trial of cognitively impaired old people referenced in Plasmalogens Parts 1, 2, and 3.
He mentions taking 5 ml in the morning and 5 ml at night because he used the Prodrome oil products. 1 ml of a Prodrome oil plasmalogen precursor product equals 900 mg of their softgel product.
“My brain is trying to minimize long-term effects of pain/stress by suppressing my memory of it. But this can only go on for so long before it becomes an entrenched state.
I have solved the sustenance side of the equation. I need to work harder to solve the environmental side.”
While I agree that we each have a responsibility to ourselves to create an environment that’s conducive to our health, the above phenomenon isn’t necessarily resolvable by changing an individual’s current environment. My understanding is that long-term effects of pain, stress, and related human experiences are usually symptoms of causes that started much earlier in our lives.
Adjusting one’s present environment may have immediate results, but probably won’t have much therapeutic impact on long-term issues. Early life memories and experiences are where we have to gradually go in order to stop being driven by what happened back then.
See Dr. Arthur Janov’s Primal Therapy for its principles and explanations. I started Primal Therapy at a similar age, 53, and continued for three years.
The 2022 plasmalogen clinical trial mentioned in Parts 1 and 2 bypassed peroxisome metabolism of cognitively impaired people per discussion of the below diagram:
Increasing the body’s fasting state with time-restricted eating, and preventing muscle atrophy with resistance exercise, were offered as the two most important ways to improve peroxisomal function.
I didn’t find any relevant 2023 human studies (where I could access the full study) on different non-drug treatments that I was willing to do. A 2023 review outlined aspects of peroxisomes, to include a few older human studies:
“Peroxisomes are small, single-membrane-bound organelles, which are dynamic and ubiquitous. Peroxisomes directly interact with other organelles, such as endoplasmic reticulum, mitochondria, or lysosomes. Peroxisomes exert different functions in various cells through both catabolic and anabolic pathways.
The main functions of peroxisomes can be categorized as reactive oxygen species (ROS) metabolism, lipid metabolism, and ether-phospholipid biosynthesis. Peroxisomes also play important roles in inflammatory signaling and the innate immune response.”
1. Since I haven’t recently tried the two main ways to improve peroxisomal function, I’ll give them a go over the next three months:
Expect to get my feeding timeframe to within eight hours. Don’t know about making it short like 6 hours, because my first meal of the day is 35 calories of microwaved cruciferous sprouts, then I wait an hour before eating anything else.
Resistance exercise progress should be measurable, as I recorded exercises during the first ten weeks of eating broccoli sprouts every day 3.5+ years ago.
2. Don’t know that I’ll recognize any cognitive improvements to the extent I did during Week 9.
I don’t have a young brain anymore, and I’m sure some decline could be measured in memory tests. But I’m not going to become a lab rat.
There’s an occasional annoyance that’s been going on for some time, especially when I’m distracted. It happens when I think of something to do, and it somehow becomes a short-term memory that I did it, instead of going into a Things To Do queue. It’s largely self-correcting. For example, regardless of what I paid, I’ll drive back to the grocery store self-checkout to retrieve a third bag that didn’t make it home. A pink-haired employee said young people leave their paid-for groceries behind all the time. It’s usually more of a reality disconnect for me than forgetfulness, because I have a memory that I performed the action. Definitely room for improvement.
3. Don’t know that I’d see biochemical changes such as some described in Part 1. Maybe I’ll move up an annual physical to compare it with the last one in May?
I already have very little oxidative stress, very little inflammation, low triglycerides, high HDL, and no major improvements are indicated on CBC / CMP / lipid panels.
Take supplements to ensure other things like acetylcholine neurotransmitter availability, one-carbon / methylation metabolism, vitamin / mineral adequacy.
4. I started the two Prodrome plasmalogen precursor supplements (ProdromeGlia and ProdromeNeuro) a week ago, and take their standard doses. My thought is that resultant plasmalogens won’t degrade very much if their primary use isn’t to immediately address oxidative stress and inflammation. That could give these extra plasmalogens a chance to make larger homeostatic contributions in myelin and membrane areas.
I don’t expect any particular effects to manifest. But I’m interested to see if these two areas would be affected:
My left ulnar nerve has been giving me problems for over five years, and several resistance exercises aggravate it. I’ve had two nerve continuity tests during that time to confirm. Numbness and pain are intermittent, though.
I still take acetaminophen several times a day for other pain.
None of the above treatments are specifically indicated. But if time-restricted feeding and/or extra plasmalogens have an effect on left ulnar or other pain, maybe I’ll be able to make better progress on resistance exercise.
Four 2023 papers that outlined or used different ergothioneine doses, starting with a human/rodent study:
“We found that cognitive function and hippocampal neurogenesis were lower in mice fed an ERGO-free diet than in those fed the control diet. Mice fed an ERGO-free diet were orally administered ERGO (0, 2, and 20 mg/kg) for two weeks which reversed these effects.
Phosphorylated brain-derived neurotrophic factor receptor TrkB, the activated form of TrkB, was also detected in extracellular vesicles (EVs) derived from serum samples of 52 volunteers who had been orally administered ERGO-containing tablets (5 mg/day for 12 weeks). The ratio of serum EV-derived phosphorylated TrkB was significantly higher in the ERGO-treated group than in the placebo-treated group and was positively correlated with both serum ERGO concentrations and several cognitive domain scores from Cognitrax.
The ratio of p-TrkB to TrkB in serum EVs was proposed as a quantitative diagnostic marker of long-term ERGO-induced cognitive improvement.”
Human equivalents of all rodent ergothioneine doses were higher than the 5 mg/day for 12 weeks 2020 human study, cited as Reference 21. I couldn’t access that paper, so here’s its Abstract:
“These results indicate that continuous intake of ergothioneine improves cognitive function in healthy subjects.”
A rodent study compared effects of a fermented product with 0.1 and 1.0 mg/g (human equivalent 6 mg (1 mg x .081) x 70 kg) ergothioneine doses:
“Our present study demonstrated for the first time the preventive effect of Rice-koji fermented extracts made by Aspergillus oryzae on anxiety, impaired recognition, and nociception using a psychophysically stressed model. Our results also demonstrated preventive effects of ergothioneine (EGT) on stress-induced anxiety- and pain-like behaviors.
Daily administration of High dose Rice-koji or 0.1 mg/kg EGT decreased anxiety- and pain-like behaviors. These findings suggest that inhibitory effects of Rice-koji on psychological stress might be mediated through the actions of EGT.”
Here’s one of several reviews that cited a 2017 clinical trial (duplicately Reference 39 and 61 for some reason) of 5 and 25 mg ergothioneine doses:
“In this pharmacokinetic study, forty-five healthy humans received placebo, 5, or 25 mg encapsulated ergothioneine/d for 7 d and were followed up for an additional 4 weeks. Ergothioneine was rapidly absorbed and largely retained by the body, with large increases in plasma ergothioneine levels and only minimal increases (<4 %) in urinary excretion observed. While plasma levels of ergothioneine decreased when supplementation was withdrawn, levels in whole blood continued to increase in a dose–response fashion, reaching maximal levels 3 weeks after withdrawal of supplement, which were sustained at 4 weeks follow-up.
A large difference in basal concentrations of ergothioneine in whole blood was observed. Participants with the highest basal levels of ergothioneine also appeared to take up more of supplemented ergothioneine.”
Wrapping up with a deep dive into seven mushroom varieties’ compounds:
“Mushrooms contain multiple essential nutrients and health-promoting bioactive compounds, including amino acid L-ergothioneine. We compared metabolomes of fresh raw white button, crimini, portabella, lion’s mane, maitake, oyster, and shiitake mushrooms using untargeted liquid chromatography mass spectrometry (LC/MS)-based metabolomics.
Results indicate significantly higher concentrations of L-ergothioneine in lion’s mane and oyster mushrooms compared to the remaining five mushroom varieties, which had concentrations ranging from 1.94 ± 0.55 to 5.26 ± 1.23 mg/100 g wet weight (mean ± SD). There was also variability in concentration of L-ergothioneine between mushroom varieties of the same farm. Different numbers denote significance (p < 0.05).
Mushrooms and their bioactive extracts are considered functional foods. Mushrooms have several bioactive compounds, including polysaccharides, lectins, terpenoids, sterols, and alkaloids, among others, which may positively impact health.
Cell walls of mushrooms contain polysaccharides, including β-glucans and chitin, which positively affect health, through modulating the immune system and protecting the cardiovascular system through improvements in glucose and lipid metabolism. Effects on the cardiovascular system are also attributable to lovastatin and polyphenols, known for their lipid-lowering and antioxidant properties, respectively.
While the 1344 compounds in common among the seven mushroom varieties support some level of similarity, detection of hundreds of unique-to-mushroom-variety compounds and differences in amino acid profiles indicate that not all mushrooms are chemically comparable. Given detection of >400 unique-to-mushroom-variety compounds in lion’s mane, maitake, oyster, and shiitake mushrooms, we suggest further targeted investigations on compounds detected and potential health benefits.”
https://www.mdpi.com/2304-8158/12/16/2985 “Metabolomics Profiling of White Button, Crimini, Portabella, Lion’s Mane, Maitake, Oyster, and Shiitake Mushrooms Using Untargeted Metabolomics and Targeted Amino Acid Analysis”
I eat around 200 grams of mushrooms daily, having temporarily overridden the boredom of eating AGE-less chicken vegetable soup every day. I prep all the top package’s frozen umami bomb (283 grams) and half of the bottom’s fresh mushrooms (340 grams) into the soup:
It makes servings for three days, including one for prep day dinner. I’d guess from “concentrations ranging from 1.94 ± 0.55 to 5.26 ± 1.23 mg/100 g (mean ± SD)” that my daily mushroom ergothioneine dose is around 7 mg ((1.94 mg + 5.26 mg) / 2) = 3.6 mg per 100 grams x 2 (for 200 grams).
This 2023 rodent study investigated relationships of MK-7 menaquinone, aluminum trichloride, and brain health:
“A variety of endogenous and exogenous agents, such as metals and environmental toxins (aluminum, mercury, etc.), can contribute to neurodegeneration, which is of multifactorial clinical occurrence.
The current study showed that Alzheimer’s Disease (AD)-like condition was induced in mice by AlCl3 treatment affecting spatial and recognition memory. Neuropathological alterations included neuroinflammation, oxidative stress, an increase in brain amyloid β levels, and loss of hippocampal neurons.
Aluminium chloride (AlCl3; 100 mg/kg for 3 weeks orally) was administered to Swiss albino mice to induce neurodegeneration and Vitamin K2 (100 mcg/kg for 3 weeks orally) was applied as treatment. This was followed by behavioral studies to determine memory changes.
Antioxidants like glutathione and SOD were low compared to the control group, while oxidative stress marker MDA was elevated. BDNF levels increased in the Vitamin K2 treated animals, suggesting its neuroprotective functions.
Vitamin K2 could partially reverse AlCl3-mediated cognitive decline. It increased hippocampal acetylcholine and BDNF levels while reducing oxidative stress, neuroinflammation, and β-amyloid deposition, protecting hippocampal neurons from AlCl3-mediated damage.“
This study’s human equivalent Vitamin K2 dose is (100 mcg x .081) x 70 kg = 567 mcg. I’ve taken 600 mcg MK-7 every day for the past two years.
Found out last week that I’ve also been inadvertently dosing myself with aluminum every day. This is the underside of my former 3-year-old drip coffee maker with its cover removed:
I’m certain its aluminum tubing that heats reservoir water started to corrode a long time ago. Currently trying out methods of making aluminum-free coffee.
“Pronounced rejuvenation effects in male rats prompted us to conduct further confirmatory experiments. A particularly important consideration is the effectiveness of E5 with regards to sex, as sex-dependent rejuvenation by some interventions have previously been reported.
To assess E5’s applicability to both male and female Sprague Dawley rats, we studied 12 males (6 treated with E5, 6 with saline) and 12 females (6 treated with E5, 6 with saline). These rats were treated every 45 days with an injection of E5 or saline. Rats were monitored for 165 days, and blood was drawn at six time points: 0, 15, 30, 60, 150 and 165 days from the first injection.
We observed highly significant improvements in TNF alpha and IL-6 levels for both males and females in the blood of E5-injected rats over that of saline controls. We also observed a substantial improvement in grip strength.
Our study shows age reversal effects in both male and female rats, but E5 is more effective in males.”
Another experimental group was started with old rats of both sexes. Using the human / rat relative clock developed in the original study, a human equivalent age to these rats at 26 months old was ((112.7 weeks / 197.6 weeks maximum rat lifespan) x 122.5 years maximum human lifespan) = 69.8 years:
“To validate our epigenetic clock results, we conducted a second set of E5 experiments with Sprague Dawley rats of both sexes. When these rats turned 26 months old, half (9 rats) received the E5 treatment while the other half (8 rats) received only the control treatment (saline injection). We analyzed methylation data from two blood draws: blood draw before treatment (baseline) and a follow up sample (15 days after the E5/saline treatment).”
Treatment measurements were affected by one female control group outlier. Panels F through J were recalculated after removing the outlier to show significant effects in both sexes:
“A) Final version of the rat clock for blood. Baseline measurement (x-axis) versus follow up measurement (15 days after treatment, y-axis). Points (rats) are colored by treatment: red=treated by E5, black=treated with saline only. Rotated grey numbers underneath each bar reports the group sizes. Each bar plot reports the mean value and one standard error.
B,D,E) Difference between follow up measurement and baseline measurement (y-axis) versus treatment status in B) all rats, D) female rats only, E) male rats only. C) is analogous to B) but uses the pan tissue clock for rats.
Panels in the second row (F,G,H,I,J) are analogous to those in the first row but the analysis omitted one control rat (corresponding to the black dot in the lower right of panel A).”
A description of how E5 plasma fraction was made starts on page 16 of the *.pdf file. The next E5 study will be done with dogs per July 2023 updates in blog post comments:
“On E5 our entire team is working hard towards the launch of an old Beagle dogs trial this month. We want to make them really young, healthy, happy, and jumping around like 1 and 2 year olds.
Primary endpoint is safety and toxicology to test various dose strengths and frequencies. Secondary endpoints are more than 20.
As you know, we like to test exhaustively to get a sharper perspective of what’s happening. In rat studies we tested 30 biomarkers, including functional. We are especially keen to check kidney markers.
There are two clocks for dogs we are interested in to get third party confirmation of age reversal. Horvath dog clock is ready and GlycanAge dog clock is under construction.
We are requesting all organizations that support pets and aging to financially support their project of building an accurate dog clock. Not only will it help veterinary aging research like ours, but also all the dog owners that may want to know how much improvement their dog received from treatment. Dr. Matt Kaeberlain is an advisor on their project.”
Three 2023 papers on the initial stage of neuronal differentiation, starting with a rodent study of taurine’s effects:
“We aimed to assess the role of taurine (TAU) in axonal sprouting against cerebral ischemic injury, clarify the function of mitochondria in TAU-induced axonal sprouting, and further determine the underlying potential molecular mechanism.
We determined that TAU improved motor function recovery and restored neurogenesis in ischemic stroke. This possibly occurred via improvements in mitochondrial function.
We investigated that the Sonic hedgehog (Shh) pathway exerted an important role in these effects. Our study findings highlighted the novel viewpoint that TAU promoted axonal sprouting by improving Shh-mediated mitochondrial function in cerebral ischemic stroke.”
A rodent study investigated effects of a soy isoflavone gut microbiota metabolite:
“Perinatally-infected adolescents living with HIV-1 (pALHIV) appear uniquely vulnerable to developing substance use disorders (SUD). Medium spiny neurons (MSNs) in the nucleus accumbens core (NAcc), an integrator of cortical and thalamic input, have been implicated as a key structural locus for the pathogenesis of SUD.
Treatment with estrogenic compounds (e.g., 17β-estradiol) induces prominent alterations to neuronal and dendritic spine structure in the NAcc supporting an innovative means to remodel neuronal circuitry. The carcinogenic nature of 17β-estradiol, however, limits its translational utility.
Plant-derived polycyclic phenols, or phytoestrogens, whose chemical structure resembles 17β-estradiol may afford an alternative strategy to target estrogen receptors. The phytoestrogen S-Equol (SE), permeates the blood-brain barrier, exhibits selective affinity for estrogen receptor β (ERβ), and serves as a neuroprotective and/or neurorestorative therapeutic for HIV-1-associated neurocognitive and affective alterations.
Beginning at approximately postnatal day (PD) 28, HIV-1 transgenic (Tg) animals were treated with a daily oral dose of 0.2 mg of SE. The SE dose of 0.2 mg was selected for two primary reasons, including:
A dose-response experimental paradigm established 0.2 mg of SE as the most effective dose for mitigating neurocognitive deficits in sustained attention in the HIV-1 Tg rat; and
The dose, which yielded a daily amount of 0.25–1.0 mg/kg/SE (i.e., approximately 2.5–10 mg in a 60 kg human), is translationally relevant (i.e., well below the daily isoflavone intake of most elderly Japanese.
Daily oral treatment continued through PD 90.
HIV-1 Tg animals exhibited an initial increase in dendrite length (A) and the number of dendritic spines (B) early in development; parameters which subsequently decreased across time. In sharp contrast, dendrite length and the number of dendritic spines were stable across development in control animals.
Targeting these alterations with the selective ERβ agonist SE during the formative period induces long-term modifications to synaptodendritic structure, whereby MSNs in the NAcc in HIV-1 Tg animals treated with SE resemble control animals at PD 180.”
A rodent brain cell study investigated soy isoflavones’ effects on a different estrogen receptor:
“We evaluated effects of isoflavones using mouse primary cerebellar culture, astrocyte-enriched culture, Neuro-2A clonal cells, and co-culture with neurons and astrocytes. Soybean isoflavone-augmented estradiol mediated dendrite arborization in Purkinje cells.
These results indicate that ERα plays an essential role in isoflavone-induced neuritogenesis. However, G-protein-coupled ER (GPER1) signaling is also necessary for astrocyte proliferation and astrocyte–neuron communication, which may lead to isoflavone-induced neuritogenesis.
We highlight the novel possibility that isoflavones enhance dendritogenesis and neuritogenesis, indicating that they can be a useful supplementary compound during brain development or in the injured brain.”