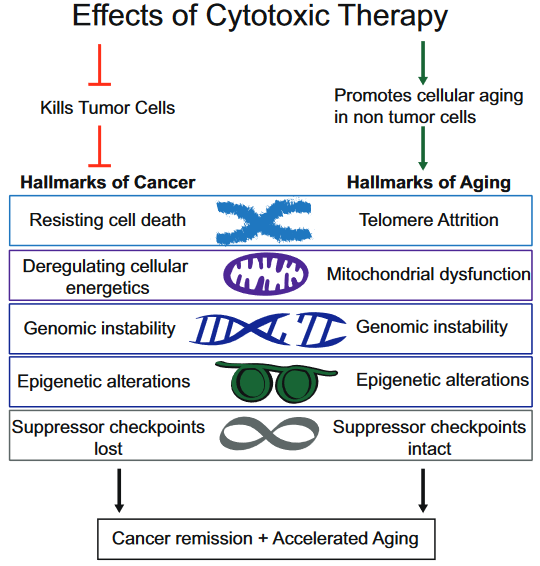
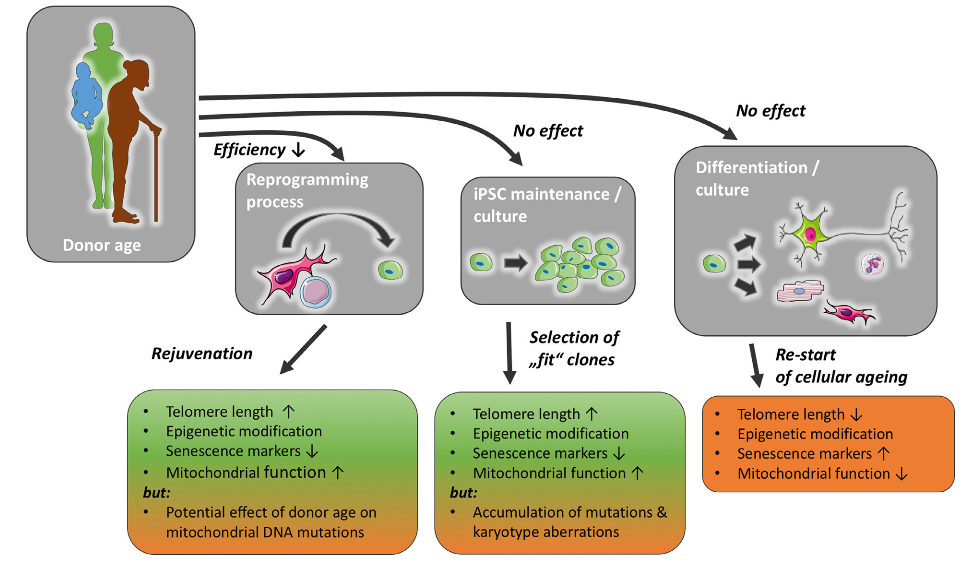
This 2018 Canadian paper reviewed evidence for potential sex-specific differences in the lasting impacts of childhood trauma:
“This paper will provide a contextualized summary of neuroendocrine, neuroimaging, and behavioral epigenetic studies on biological sex differences contributing to internalizing psychopathology, specifically posttraumatic stress disorder and depression, among adults with a history of childhood abuse.
Given the breadth of this review, we limit our definition [of] trauma to intentional and interpersonal experiences (i.e., childhood abuse and neglect) in childhood. Psychopathological outcomes within this review will be limited to commonly explored internalizing disorders, specifically PTSD and depression.
Despite the inconsistent and limited findings in this review, a critical future consideration will be whether the biological effects of early life stress can be reversed in the face of evidence-based behavioral interventions, and furthermore, whether these changes may relate to potentially concurrent reductions in susceptibility to negative mental health outcomes.”
It was refreshing to read a paper where the reviewers often interrupted the reader’s train of thought to interject contradictory evidence, and display the scientific method. For example, immediately after citing a trio of well-respected studies that found:
“Psychobiological research on relationships linking impaired HPA axis functioning and adult internalizing disorders are suggestive of lower basal and afternoon levels of plasma cortisol in PTSD phenotype.”
the reviewers stated:
“However, a recent meta-analysis suggests no association between basal cortisol with PTSD.”
and effectively ended the cortisol discussion with:
“Findings are dependent upon variance in extenuating factors, including but not limited to, different measurements of:
- early adversity,
- age of onset,
- basal cortisol levels, as well as
- trauma forms and subtypes, and
- presence and severity of psychopathology symptomology.”
The reviewers also provided good summaries of aspects of the reviewed subject. For example, the “Serotonergic system genetic research, childhood trauma and risk of psychopathology” subsection ended with:
“Going forward, studies must explore the longitudinal effects of early trauma on methylation as well as comparisons of multiple loci methylation patterns and interactions to determine the greatest factors contributing to health outcomes. Only then, can we start to consider the role of sex in moderating risk.”
I didn’t agree with the cause-ignoring approach of the behavior therapy mentioned in the review. Does it make sense to approach one category of symptoms:
“the biological effects of early life stress”
by treating another category of symptoms?
“can be reversed in the face of evidence-based behavioral interventions.”
But addressing symptoms instead of the sometimes-common causes that generate both biological and behavioral effects continues to be the direction.
After receiving short-term symptom relief, wouldn’t people prefer treatments of originating causes so that their various symptoms don’t keep bubbling up? Why wouldn’t research paradigms be aligned accordingly?
I was encouraged by the intergenerational and transgenerational focus of one of the reviewer’s research:
“Dr. Gonzalez’s current research focus is to understand the mechanisms by which early experiences are transmitted across generations and how preventive interventions may affect this transmission.”
This line of hypotheses requires detailed histories, and should uncover causes for many effects that researchers may otherwise shrug off as unexplainable individual differences. Its aims include the preconception through prenatal periods when both the largest and the largest number of epigenetic changes occur, and is when our susceptibility and sensitivity to our environment is greatest. There are fewer opportunities for effective “preventive interventions” in later life compared with these early periods.
Unlike lab rats, women and men can reach some degree of honesty about our early lives’ experiential causes of ongoing adverse effects. Experiential therapies that allow humans to potentially change their responses to these causes deserve more investigation than do therapies that apply external “interventions.”
https://www.sciencedirect.com/science/article/pii/S0272735817302647 “Biological alterations affecting risk of adult psychopathology following childhood trauma: A review of sex differences” (not freely available) Thanks to lead author Dr. Ashwini Tiwari for providing a copy.