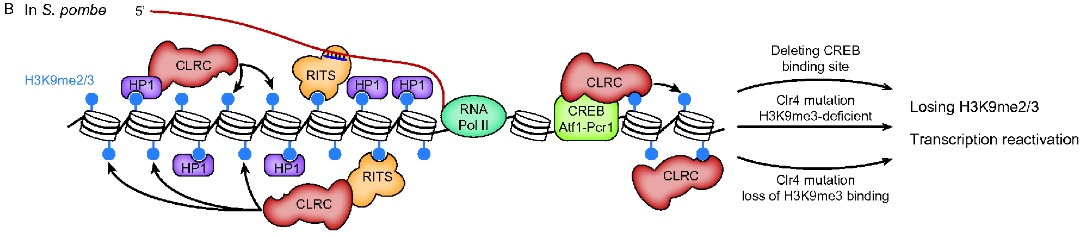
This 2018 Washington rodent study subject was transgenerational epigenetic inheritance of disease caused by a fungicide that’s been phased out or banned for over a decade:
“This study was designed to help understand how three different epigenetic processes in sperm are correlated with vinclozolin-induced epigenetic transgenerational inheritance of disease.
- Most DMRs [differential DNA-methylated regions] identified in this study are unique between the F1, F2, and F3 generations.
- The number of lncRNA was much higher than the number of sncRNA [small noncoding RNA, including microRNA]. The overlap between each generation was very low or nonexistent.
- The F1 and the F2 generation control versus vinclozolin lineage sperm had negligible DHRs [differential histone retention sites]. This observation suggests that the direct vinclozolin exposure does not alter histone retention or trigger any changes. However, the F3 generation control versus vinclozolin lineage sperm DHRs increased considerably.
It appears that the phenomenon is more complex than just a direct exposure triggering the formation of epimutations that are then simply maintained in the subsequent generations.”

There’s something odd about a study where a third of the 87 cited references list one of the study’s coauthors, who also coauthored A review of epigenetic transgenerational inheritance of reproductive disease. I couldn’t find a satisfactory explanation for the study’s over-the-top self-referencing.
What do you think?
I asked the coauthors why a third of the cited references were self-referencing. The lead author replied:
“The field in epigenetic transgenerational inheritance is expanding, however it is still hard for us to find relevant studies in rodents or human that we can cite. Most of the time DNA methylation, ncRNA and histone modifications are investigated from a direct exposure and/or from a purely mechanistic angle (e.g. DNA methylation of specific genes).
In contrast, transgenerational phenotypes and toxicology by definition excludes direct exposure and must be transmitted through multiple generations (the F3 generation is the first transgenerational one). We are not looking at specific genes but using whole genome sequencing technologies which is a broader approach.
Besides, if you do a pubmed search with the keywords “epigenetics” and “transgenerational”, you will probably find that more than 50% of the studies have been done by Dr Michael K. Skinner. He is also one of the first researcher who started to work on the epigenetic transgenerational inheritance phenomenon 15 years ago. Not citing his previous work is challenging.
We hope to see other labs contributing to this particular field and we will be delighted to cite them. In the meantime, our only option is to reference our previous work.”
I replied:
“Thank you for your reply! It must be exasperating to see other researchers stop their studies short of the F3 generation for no apparent or disclosed reason.
Have you seen even one scientifically adequate human study of transgenerational epigenetic inheritance?”
https://academic.oup.com/eep/article/4/2/dvy010/4987173 “Alterations in sperm DNA methylation, non-coding RNA expression, and histone retention mediate vinclozolin-induced epigenetic transgenerational inheritance of disease”