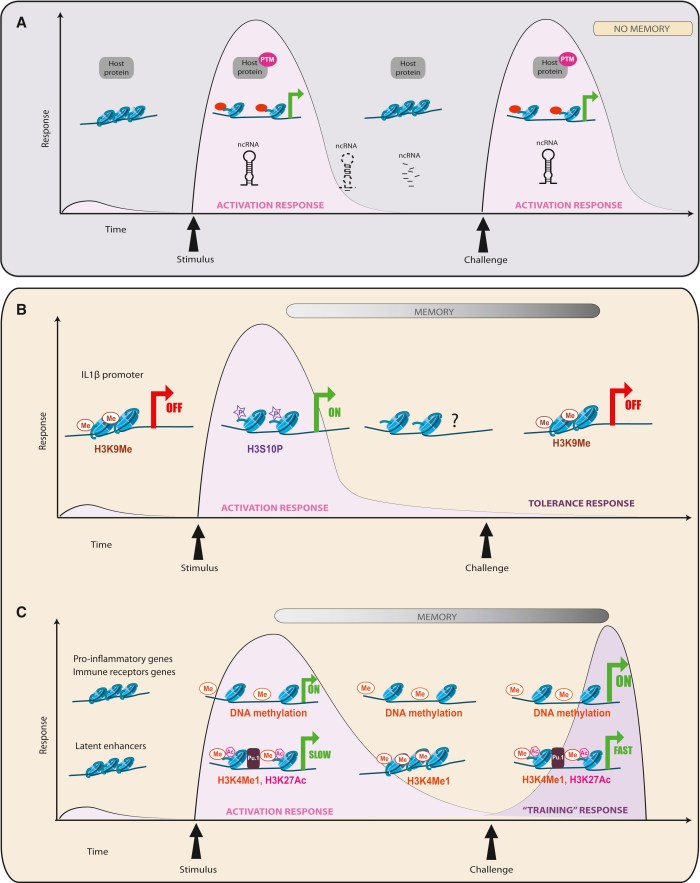
This 2017 Netherlands review subject was the lasting epigenetic effects of early-life stress:
“Exposure to stress during critical periods in development can have severe long-term consequences.
One of the key stress response systems mediating these long-term effects of stress is the hypothalamic-pituitary-adrenal (HPA) axis.
Early life stress (ELS) exposure has been reported to have numerous consequences on HPA-axis function in adulthood.
ELS is able to “imprint” or “program” an organism’s neuroendocrine, neural and behavioral responses to stress. Research focuses along two complementary lines:
- ELS during critical stages in brain maturation may disrupt specific developmental processes (by altered neurotransmitter exposure, gene transcription, or neuronal differentiation), leading to aberrant neural circuit function throughout life.
- ELS may induce modifications of the epigenome which lastingly affect brain function.
These epigenetic modifications are inducible, stable, and yet reversible, constituting an important emerging mechanism by which transient environmental stimuli can induce persistent changes in gene expression and ultimately behavior.”
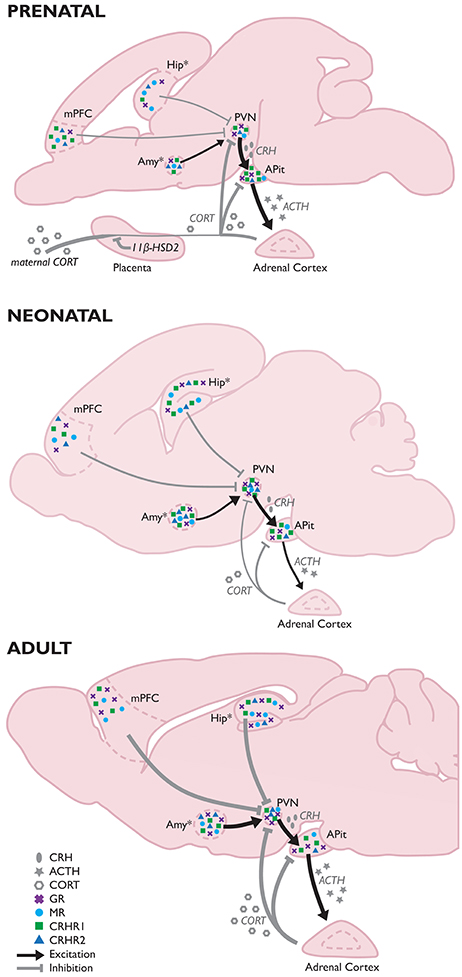
In early life, the lower brain and limbic system brain structures are more developed and dominant, whereas the cerebrum is less developed (use the above rodent graphic as a rough guide). Stress and pain generally have a greater impact on a fetus than an infant, and a greater impact on an infant than an adult.
The reviewers cited 50+ studies from years 2000-2015 in the “Early Life Stress Effects in a “Matching” Stressful Adult Environment” section to argue for the match / mismatch theory:
“Encountering ELS prepares an organism for similar (“matching”) adversities during adulthood, while a mismatching environment results in an increased susceptibility to psychopathology, indicating that ELS can exert either beneficial or disadvantageous effects depending on the environmental context.
Initial evidence for HPA-axis hypo-reactivity is observed for early social deprivation, potentially reflecting the abnormal HPA-axis function as observed in post-traumatic stress disorder.
Experiencing additional (chronic) stress in adulthood seems to normalize these alterations in HPA-axis function, supporting the match / mismatch theory.”
Evidence for this theory was contrasted with the allostatic load theory presented in How one person’s paradigms regarding stress and epigenetics impedes relevant research.
The review mainly cited evidence from rodent studies that mismatched reactions in adulthood may be consequences of early-life events. These events:
“Imprint or program an organism’s neuroendocrine, neural and behavioral responses..leading to aberrant neural circuit function throughout life..which lastingly affect brain function.”
Taking this research to a personal level:
- Have you had feelings that you were unsafe, although your environment was objectively safe?
- Have you felt uneasy when people are nice to you?
- Have you felt anxious when someone pays attention to you, even after you’ve acted to gain their attention?
Mismatched human feelings are one form of mismatched reactions. These may be consequences of early-life experiences, and indicators of personal truths.
If researchers can let go of their biases and Advance science by including emotion in research, they may find that human subjects’ feelings produce better evidence for what actually happened during the subjects’ early lives than do standard scientific methods of:
- Surveys of the responsible parties (for example, Parental lying thwarted both their children and researchers); and
- Self-reports of early-life events (for example, A problematic study of oxytocin receptor gene methylation, childhood abuse, and psychiatric symptoms).
Incorporating feeling evidence may bring researchers and each individual closer to discovering the major insults that knocked their development processes out of normally robust pathways and/or induced “persistent changes in gene expression and ultimately behavior.”
https://www.frontiersin.org/articles/10.3389/fncel.2017.00087/full “Modulation of the Hypothalamic-Pituitary-Adrenal Axis by Early Life Stress Exposure”
I came across this review as a result of it being cited in http://www.sciencedirect.com/science/article/pii/S1084952117302884 “Long-term effects of early environment on the brain: Lesson from rodent models” (not freely available)