People will forgive you for being wrong, but they will never forgive you for being right – especially if events prove you right while proving them wrong. Thomas Sowell
Home* rating2-3 stars Required further work(Page 11)
“A single dimension is able to measure a person’s liability to mental disorder, comorbidity among disorders, persistence of disorders over time, and severity of symptoms.”
The coauthors partially based this on:
“Repeated diagnostic interviews carried out over 25 years, when the research participants were 11, 13, 15, 18, 21, 26, 32, and 38 years old, and include information about seven diagnostic groups: anxiety, depression, attention deficit hyperactivity disorder, conduct disorder, substance dependence, bipolar disorder, and schizophrenia.”
“Dunedin and other studies show that most people have at least one episode of mental illness during their lifetime.”
What compels people to manufacture “universal” truths? Aren’t such beliefs poor substitutes for feeling? For understanding historical, factual, personal truths?
What if the price we pay for avoiding and pressuring down our feelings is: A wasted life?
What if the grand hypothesis worth proving is: For one’s life to have meaning, each individual has to regain their feelings?
The founder of the epigenetic clock technique was interviewed for MIT Technology Review:
“We need to find ways to keep people healthier longer,” he says. He hopes that refinements to his clock will soon make it precise enough to reflect changes in lifestyle and behavior.”
The journalist attempted to dumb the subject down “for the rest of us” with distortions such as the headline. The varying correlation of epigenetic age to chronological age was somewhat better reported in the story:
“The epigenetic clock is more accurate the younger a person is. It’s especially inaccurate for the very old.”
The journalist inappropriately used luck as a synonym for randomness/stochasticity:
“He estimates that about 40% of the ticking rate is determined by genetic inheritance, and the rest by lifestyle and luck.”
A third example of less-than-straightforward journalism started with:
“Such personalization raises questions about fairness. If your epigenetic clock is ticking faster through no fault of your own..”
Were MIT Technology Review readers unable to comprehend a straightforward story on the epigenetic clock? What was the purpose of slants and distortions in an introductory article?
A subset of memory recall–induced neurons in the DG [dentate gyrus] becomes reactivated after memory attenuation,
The degree of fear reduction positively correlates with this reactivation, and
The continued activity of memory recall–induced neurons is critical for remote fear memory attenuation.
Although other brain areas such as the prefrontal cortex and the amygdala are likely to be implicated in remote fear memories and remain to be investigated, these results suggest that fear attenuation at least partially occurs in memory recall–induced ensembles through updating or unlearning of the original memory trace of fear.
These data thereby provide the first evidence at an engram-specific level that fear attenuation may not be driven only by extinction learning, that is, by an inhibitory memory trace different from the original fear trace.
Rather, our findings indicate that during remote fear memory attenuation both mechanisms likely coexist, albeit with the importance of the continued activity of memory recall–induced neurons experimentally documented herein. Such activity may not only represent the capacity for a valence change in DG engram cells but also be a prerequisite for memory reconsolidation, namely, an opportunity for learning inside the original memory trace.
As such, this activity likely constitutes a physiological correlate sine qua non for effective exposure therapies against traumatic memories in humans: the engagement, rather than the suppression, of the original trauma.”
The researchers also provided examples of human trauma:
“We dedicate this work to O.K.’s father, Mohamed Salah El-Dien, and J.G.’s mother, Wilma, who both sadly passed away during its completion.”
So, how can this study help humans? The study had disclosed and undisclosed limitations:
1. Humans aren’t lab rats. We can ourselves individually change our responses to experiential causes of ongoing adverse effects. Standard methodologies can only apply external treatments.
2. It’s a bridge too far to go from neural activity in transgenic mice to expressing unfounded opinions on:
“A physiological correlate sine qua non for effective exposure therapies against traumatic memories in humans.”
Human exposure therapies have many drawbacks, in addition to being applied externally to the patient on someone else’s schedule. A few others were discussed in The role of DNMT3a in fear memories:
“Inability to generalize its efficacy over time,
Potential return of adverse memory in the new/novel contexts,
Context-dependent nature of extinction which is widely viewed as the biological basis of exposure therapy.”
3. Rodent neural activity also doesn’t elevate recall to become an important goal of effective human therapies. Clearly, what the rodents experienced should have been translated into human reliving/re-experiencing, not recall! Terminology used in animal studies preferentially has the same meaning with humans, since the purpose of animal studies is to help humans.
4. The researchers acknowledged that:
“Other brain areas such as the prefrontal cortex and the amygdala are likely to be implicated in remote fear memories and remain to be investigated.”
“The findings imply that in response to traumatic stress, some individuals, instead of activating the glutamate system to store memories, activate the extra-synaptic GABA system and form inaccessible traumatic memories.”
The study I curated yesterday, Organ epigenetic memory, demonstrated organ memory storage. It’s hard to completely rule out that other body areas may also store traumatic memories.
The wide range of epigenetic memory storage vehicles is one reason why effective human therapies need to address the whole person, the whole body, and each individual’s entire history.
This post has somehow become a target for spammers, and I’ve disabled comments. Readers can comment on other posts and indicate that they want their comment to apply here, and I’ll re-enable comments.
This 2017 UC Irvine human review subject provided details of how fetalhypothalamic-pituitary-adrenal components and systems develop, and how they are epigenetically changed by the mother’s environment:
“The developmental origins of disease or fetal programming model predicts that intrauterine exposures have life-long consequences for physical and psychological health. Prenatal programming of the fetal hypothalamic-pituitary-adrenal (HPA) axis is proposed as a primary mechanism by which early experiences are linked to later disease risk.
Development of the fetal HPA axis is determined by an intricately timed cascade of endocrine events during gestation and is regulated by an integrated maternal-placental-fetal steroidogenic unit. Mechanisms by which stress-induced elevations in hormones of maternal, fetal, or placental origin influence the structure and function of the emerging fetal HPA axis are discussed.
Human gestational physiology and fetal HPA axis development differ even from that of closely related nonhuman primates, thereby limiting the generalizability of animal models. This review will focus solely on studies of prenatal stress and fetal HPA axis development in humans.”
1. Every time I read a prenatal study I’m in awe of all that has to go right – and at the appropriate times and sequences – for a fetus to be undamaged. Add in what needs to happen at birth, during infancy, and throughout early childhood, and it seems impossible for any human to escape epigenetic damage.
2. The reviewers referenced animal studies and human research performed with postnatal subjects, despite the disclaimer:
This review will focus solely on studies of prenatal stress and fetal HPA axis development in humans.”
This led to blurring of what had been studied or not with human fetuses regarding the subject.
3. These reviewers uncritically listed many dubious human studies that had both stated and undisclosed severe limitations on their findings. Other reviewers offer informed analysis of cited studies, as Sex-specific impacts of childhood trauma summarized with cortisol:
“Findings are dependent upon variance in extenuating factors, including but not limited to, different measurements of:
presence and severity of psychopathology symptomology.”
4. The paper would have been better had it stayed on topic with its title “Developmental origins of the human hypothalamic-pituitary-adrenal axis.” Let other reviews cover animals, post-natal humans, and questionable evidence.
5. I asked the reviewers to provide a searchable file to facilitate using their work as a reference.
This 2018 Chinese study electronically modeled the brain’s circuits to evaluate memory transfer mechanisms:
“During non-rapid-eye-movement (NREM) sleep, thalamo-cortical spindles and hippocampal sharp wave-ripples have been implicated in declarative memory consolidation. Evidence suggests that long-term memory consolidation is coordinated by the generation of:
enabling memory transfer from the hippocampus to the cortex.
Consolidation has also been demonstrated in other brain tasks, such as:
In the acquisition of motor skills, where there is a shift from activity in prefrontal cortex to premotor, posterior parietal, and cerebellar structures; and
In the transfer of conscious to unconscious tasks, where activity in initial unskilled tasks and activity in skilled performance are located in different regions, the so-called ‘scaffolding-storage’ framework.
By separating a neural circuit into a feedforward chain of gating populations and a second chain coupled to the gating chain (graded chain), graded information (i.e. information encoded in firing rate amplitudes) may be faithfully propagated and processed as it flows through the circuit. The neural populations in the gating chain generate pulses, which push populations in the graded chain above threshold, thus allowing information to flow in the graded chain.
In this paper, we will describe how a set of previously learned synapses may in turn be copied to another module with a pulse-gated transmission paradigm that operates internally to the circuit and is independent of the learning process.”
The study had neither been peer-reviewed, nor were the mechanisms tested in living beings.
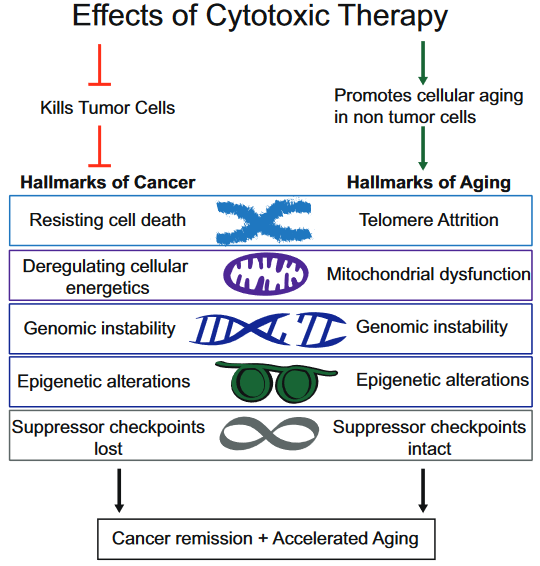
This 2018 UC San Diego review subject was the interplay between breast cancer treatments and their effects on aging:
“Although current breast cancer treatments are largely successful in producing cancer remission and extending lifespan, there is concern that these treatments may have long lasting detrimental effects on cancer survivors, in part, through their impact on non-tumor cells. It is unclear whether breast cancer and/or its treatments are associated with an accelerated aging phenotype.
In this review, we have highlighted five of nine previously described cellular hallmarks of aging that have been described in the context of cytotoxic breast cancer treatments:
The review was full of caveats weakening the above graphic’s associations:
“Telomere attrition – Blood TL [telomere length] was not associated with chemotherapy in three out of four studies;
Mitochondrial dysfunction – How cancer therapies affect cellular energetics as they relate to rate of aging is unclear;
Genomic instability – Potentially contributing to accelerated aging;
Epigenetic alterations – Although some of the key regulators of these processes have begun to be identified, including DNA and histone methylases and demethylases, histone acetylases and de-acetylases and chromatin remodelers, how they regulate the changes in aging through alteration of global transcriptional programs, remains to be elucidated; and
Cellular senescence – Dysregulated pathways can be targeted by cytotoxic chemotherapies, resulting in preferential cell death of tumor cells, but how these treatments also affect normal cells with intact pathways is unclear.”
To their credit, these reviewers at least presented some of the contrary evidence, and didn’t continue on with a directed narrative as other reviewers are prone to do.
The originator of the epigenetic clock methodology was a coauthor of the review. Only one of his works was cited in the Epigenetic alterations subsection:
This freely-available 2017 study quoted below highlighted that epigenetic clock measurements as originally designed were tissue-specific:
“To our knowledge, this is the first study to demonstrate that breast tissue epigenetic age exceeds that of blood tissue in healthy female donors. In addition to validating our earlier finding of age elevation in breast tissue, we further demonstrate that the magnitude of the difference between epigenetic age of breast and blood is highest in the youngest women in our study (age 20–30 years) and gradually diminishes with advancing age. As women approach the age of the menopausal transition, we found that the epigenetic of age of blood approaches that of the breast.”
Additional caution was justified in both interpreting age measurements and extending them into “cellular hallmarks” when the tissue contained varying cell types:
“Our studies were performed on whole breast tissue. Diverse types of cells make up whole breast tissue, with the majority of cells being adipocytes. Other types of cells include epithelial cells, cuboidal cells, myoepithelial cells, fibroblasts, inflammatory cells, vascular endothelial cells, preadipocytes, and adipose tissue macrophages.
This raises the possibility that the magnitude of the effects we observe, of breast tissue DNAm age being greater than other tissues, might be an underestimation, since it is possible that not all of the cells of the heterogenous sample have experienced this effect. Since it is difficult to extract DNA from adipose tissue, we suspect that the majority of DNA extracted from our whole breast tissues was from epithelial and myoepithelial cells.”
This 2018 German review provided short summaries of 44 studies on the contribution of histone modifications to allergies. An overall summary of their search results was:
“There are at least two levels at which the role of histone modifications is manifested.
One is the regulation of cells that contribute to the allergic inflammation (T cells and macrophages) and those that participate in airway remodeling.
The other is the direct association between histone modifications and allergic phenotypes.
Inhibitors of histone-modifying enzymes may potentially be used as anti-allergic drugs. Furthermore, epigenetic patterns may provide novel tools in the diagnosis of allergic disorders.”
This type of search is what’s expected of researchers who will perform either:
A meta-analysis of studies selected from the search results; or
Their own study.
These reviewers didn’t indicate that they were proceeding along either path.
The review was fine for the purpose of presenting current studies of the subject. But this was just the preparatory stage of research.
This 2018 German review was comprehensive for its subject, epigenetic control of variation and stochasticity in metabolic disease. I’ll focus on one aspect, phenotypic variation:
“Phenotypic [Mendelian] variation can result both from gain- and loss-of-function mutations. Because of the extreme interconnectivity of cell regulatory networks, even at the cellular level, predicting the impact of a sequence variant is difficult as the resultant variation acts:
In the context of all other variants and
Their potential additive, synergistic and antagonistic interactions.
This phenomenon is known as epistasis.
∼98.5% of our genome is non-protein-coding: it is pervasively transcribed, and its transcripts can support regulatory function. Among the best functionally characterized non-coding RNAs (ncRNAs) arising from these sequences are microRNAs (miRNAs).
Environmental [non-Mendelian] variation or ‘stimuli’ occurring during critical windows of susceptibility can elicit lifelong alterations in an individual’s phenotype. Intergenerational metabolic reprogramming [in fruit flies] results from global alterations in chromatin state integrity, particularly from reduced H3K27me3 and H3K9me3 [histone] domains.
The broad variation of fingerprints in humans is thought to depend to a large degree on stochastic variation in mechanical forces. These clear examples of inducible multi-stable or stochastic variation highlight how little we know about the landscape of potential phenotypic variation itself.
Consensus estimates of heritability for obesity and T2D are ∼70% and ∼35% respectively. The remaining, unexplained component is known to involve gene–environment interactions as well as non-Mendelian players.”
Although the above graphic displays transgenerational inheritance for humans, the reviewers didn’t cite any human studies that adequately demonstrated causes for and effects of transgenerational epigenetic inheritance.
I’ve read the cited Swedish and Dutch studies. Their designs, methods, and “correlate with” / “was associated with” results didn’t provide incontrovertible evidence from the F0 great-grandparents, F1 grandparents, F2 parents, and F3 children. It’s necessary to thoroughly study each generation to confirm definitive transgenerational epigenetic inheritance causes and effects.
As noted in How to hijack science: Ignore its intent and focus on the 0.0001%, there aren’t any such published studies to cite. Researchers urgently need to do this human research, and stop using these poor substitutes [1] to pretend there are already adequately evidenced transgenerational epigenetic inheritance human results.
I downgraded the review for treating research of this and other subjects as faits accomplis. It’s opposite ends of the evidential spectrum to state “how little we know about the landscape of potential phenotypic variation,” and in the same review, speciously extrapolate animal experiments into putative human results.
[1] As an example of the poor substitutes for evidence, a researcher referred me to the 2013 “Transgenerational effects of prenatal exposure to the 1944–45 Dutch famine” which is freely available at https://obgyn.onlinelibrary.wiley.com/doi/full/10.1111/1471-0528.12136 as a study finding human transgenerational epigenetic inheritance.
The Methods section showed:
The study’s non-statistical data was almost all unverified self-reports by a self-selected sample of the F2 generation, average age 37.
No detailed physical measurements or samples were taken of them, nor of the F1 generation, nor of the F0 generation, all of which are required as baselines for any transgenerational epigenetic inheritance findings.
No detailed physical measurements or samples were taken of the F3 generation, which is the generation that may provide transgenerational evidence if the previous generations also have detailed physical baselines.
The study’s researchers drew enough participants (360) such that their statistics package allowed them to impute and assume into existence a LOT of data. But the scientific method constrained them to make factual statements of what the evidence actually showed. They admitted:
“In conclusion, we did not find a transgenerational effect of prenatal famine exposure on the health of grandchildren in this study.”
Yet this study is somehow cited for evidence of human transgenerational epigenetically inherited causes and effects!
I’m curating this 2018 UC Berkeley/Drexel/Netherlands analysis of human studies via its press coverage. The authors:
“Collaborated to analyze data on hundreds of adults – some mentally or physically sound, others suffering from various conditions such as depression, anxiety, or post-traumatic stress disorder. Participants had completed surveys about their mental health and had their heart rates monitored via electrocardiogram.
Researchers used the data to conduct six different experiments. They sought to find out whether the conclusions of each study would successfully apply to participants individually.
One study that focused on how frequently depression sufferers reported feeling worried. Results tallied from the pool of participants showed that depressed people worry a significant amount.
But when the analysis was applied individually, the results were all over the map. Some participants worried hardly at all, while others were notably beyond the group average.
Another experiment that centered around the link between fear and avoidance showed a strong correlation when measured as a group. Yet a significant number of participants who experienced fear had no issues with avoiding various activities.
Across all six experiments, the authors could not show that what was concluded for the group applied to most individuals.”
“The probability of turning right averaged across all individuals within each line was statistically indistinguishable from 50%. However, an individual fly’s probability of turning right often diverged markedly from the population average.”
“Epigenomic analyses are limited by averaging of population-wide dynamics and do not inform behavior of single cells. We observe dynamics at the single-cell level not predicted by epigenomic analysis.”
“It is critical to obtain a measurement of variability that is independent of the mean to ensure to not confound changes in variability with shifts in mean.”
“16 of the 18 individual age acceleration estimates of the control group subjects aren’t close to the median value!”
The current study highlighted the fact that people aren’t interchangeable. Assuming ergodicity is a statistical analysis flaw that produces individually inapplicable results for many measurements of fruit flies, cells, humans, you name the organism.
This 2018 Korean review discussed aspects of the hypothalamus and aging:
“A majority of physiological functions that decline with aging are broadly governed by the hypothalamus, a brain region controlling development, metabolism, reproduction, circadian rhythm, and homeostasis. In addition, the hypothalamus is poised to connect the brain and the body so that the environmental information affecting aging can be transmitted through the hypothalamus to affect the systematic aging of the peripheral organs.
The hypothalamus is hypothesized to be a primary regulator of the process of aging of the entire body. This review aims to assess the contribution of hypothalamic aging to the age-related decline in body functions, particularly from the perspective of:
energy homeostasis,
hormonal balance,
circadian rhythm, and
reproduction,
and to highlight its underlying cellular mechanisms with a focus on:
The reviewers didn’t consider aging to be an “unintended consequence” of development. This perspective was found in a reference to A study of DNA methylation and age:
“Aging is not programmed. Instead, aging is a continuation of developmental growth, driven by genetic pathways.
Genetic programs determine developmental growth and the onset of reproduction. When these programs are completed, they are not switched off.
Aging has no purpose (neither for individuals nor for group), no intention. Nature does not select for quasi-programs. It selects for robust developmental growth.”
“The proposed epigenetic clock theory of ageing views biological ageing as an unintended consequence of both developmental programmes and maintenance programmes.”
“The hypothalamus is hypothesized to be a primary regulator of the process of aging.”
Almost all of the details discussed were from rodent studies.
As detailed in How to cure the ultimate causes of migraines? and its references, the hypothalamus is a brain structure that lacks feedback mechanisms for several of its activities. This structure develops shortly after conception and has an active prenatal role.
The hypothalamus plays its part in getting us developed and ready to reproduce, with certain feedback loops being evolutionarily unnecessary. The hypothalamus perfectly illustrates the point of:
“When these programs are completed, they are not switched off.”
Evolutionarily unnecessary feedback for aspects of hypothalamic activity may result in it not winding down when its developmental role is over. This activity shouldn’t be interpreted to construe a role that has some other meaning or purpose.
This 2018 Michigan review subject was cancer evolution:
“Based on the fact that cancer typically represents a complex adaptive system, where there is no linear relationship between lower-level agents (such as each individual gene mutation) and emergent properties (such as cancer phenotypes), we call for a new strategy based on the evolutionary mechanism of aneuploidy [abnormal number of chromosomes] in cancer, rather than continuous analysis of various individual molecular mechanisms.
Cancer evolution can be understood by the dynamic interaction among four key components:
Internal and external stress;
Elevated genetic and non-genetic variations (either necessary for cellular adaptation or resulting from cellular damages under stress);
Genome-based macro-cellular evolution (genome replacement, emergent as new systems); and
Multiple levels of system constraint which prevent/slow down cancer evolution (from tissue/organ organization to the immune system interaction).
Since the sources of stress are unlimited and unavoidable (as they are required by all living systems), there are large numbers of gene mutations / epigenetic events / chromosomal aberrations, such as aneuploidy, that can be linked to stress-mediated genomic variants. Furthermore, as environmental constraints are constantly changing, even identical instances of aneuploidy will have completely different outcomes in the context of cancer evolution, as the results of each independent run of evolution will most likely differ.
Most current research efforts are focusing on molecular profiles based on an average population, and outliers are eliminated or ignored, either by the methods used or statistical tools. The traditional view of biological research is to identify patterns from “noise,” without the realization that the so-called “noise” in fact is heterogeneity, which represents a key feature of cancer evolution by functioning as the evolutionary potential.
Understanding the molecular mechanism (both cause and effect) of aneuploidy is far from enough. A better strategy is to monitor the evolutionary process by measuring evolutionary potential. For example, the overall degree of CIN [chromosome instability] is more predictive than individual gene mutation profile.”
Although I read many abstracts of cancer research papers every week, I usually don’t curate them. I curated this paper because the reviewers emphasized several themes of this blog, including:
Further examples of how stress may shape one’s life.
How researchers miss information when they ignore or process away variation:
“Studies have demonstrated the importance of outliers in cancer evolution, as cancer is an evolutionary game of outliers. While this phenomenon can provide a potential advantage for cellular adaptation, it can also, paradoxically, generate non-specific system stress, which can further produce more genetic and non-genetic variants which favor the disease condition.”
Epigenetics researchers may benefit from evolutionary viewpoints that incorporate the interactions of stress and “genetic and non-genetic variants.”
Since epigenetic changes require inheritance in order to persist, it would be a step forward to see researchers start “measuring evolutionary potential” of these inheritance processes.
This 2018 Israeli human study subject was natural killer cell epigenetic memory of pregnancies:
“Natural killer (NK) cells were first discovered for their ability to kill tumor cells, and later found to also kill pathogen-infected cells.
Different tissue-resident subpopulations of human NK cells exist throughout the body, displaying unique phenotypic and functional properties. One of the most fascinating tissue-resident subsets of NK cells, termed decidual NK cells, is found at the maternal fetal interface (decidua) in direct contact with the placenta.
We discovered a population found in repeated pregnancies, which has a unique transcriptome and epigenetic signature..have open chromatin around the enhancers of [growth factor genes] IFNG [essential for angiogenesis] and VEGFA [supporting vascular formation].
The pregnancy-related NK memory cells identified here might represent the first example of improved function of NK cells that occurs under healthy physiological conditions.”
One source for the experiments was:
“Decidual samples from healthy women who underwent elective first trimester terminations of normal pregnancies.”
We’ll start with a 2018 epigenetic clock human study from Finland:
“We evaluated the association between maternal antenatal depression and a novel biomarker of aging at birth, namely epigenetic gestational age (GA) based on fetal cord blood methylation data. We also examined whether this biomarker prospectively predicts and mediates maternal effects on early childhood psychiatric problems.
Maternal history of depression diagnosed before pregnancy and greater antenatal depressive symptoms were associated with child’s lower epigenetic GA. Child’s lower epigenetic GA, in turn, prospectively predicted total and internalizing problems and partially mediated the effects of maternal antenatal depression on internalizing problems in boys.”
Listening to a podcast by one of the coauthors, although the researchers’ stated intent was to determine the etiology of the findings, I didn’t hear any efforts to study the parents in sufficient detail to be able to detect possible intergenerational and transgenerational epigenetic inheritance causes and effects. There were the usual “associated with” and “it could be this, it could be that” hedges, which were also indicators of the limited methods employed toward the study’s limited design.
Why was an opportunity missed to advance human research in this area? Are researchers satisfied with non-causal individual differences non-explanations instead of making efforts in areas that may produce etiological findings?
The second 2018 epigenetic clock human study was from Alabama:
“We estimated measures of epigenetic age acceleration in 830 Caucasian participants from the Genetics Of Lipid Lowering Drugs and diet Network (GOLDN) considering two epigenetic age calculations.
Both DNA methylation age estimates were highly correlated with chronological age. We found that the Horvath and Hannum measures of epigenetic age acceleration were moderately correlated.
The Horvath age acceleration measure exhibited marginal associations with increased postprandial [after eating a meal] HDL [high-density lipoprotein], increased postprandial total cholesterol, and decreased soluble interleukin 2 receptor subunit alpha (IL2sRα). The Hannum measure of epigenetic age acceleration was inversely associated with fasting HDL and positively associated with postprandial TG [triglyceride], interleukin-6 (IL-6), C-reactive protein (CRP), and tumor necrosis factor alpha (TNFα).
The third 2018 epigenetic clock human study was a meta-analysis of cohorts from the UK, Italy, Sweden, and Scotland:
“The trajectories of Δage showed a declining trend in almost all of the cohorts with adult sample collections. This indicates that epigenetic age increases at a slower rate than chronological age, especially in the oldest population.
Some of the effect is likely driven by survival bias, where healthy individuals are those maintained within a longitudinal study, although other factors like underlying training population for the respective clocks may also have influenced this trend. It may also be possible that there is a ceiling effect for Δage whereby epigenetic clock estimates plateau.”
This 2018 Washington rodent study subject was transgenerational epigenetic inheritance of disease caused by a fungicide that’s been phased out or banned for over a decade:
“This study was designed to help understand how three different epigenetic processes in sperm are correlated with vinclozolin-induced epigenetic transgenerational inheritance of disease.
Most DMRs [differential DNA-methylated regions] identified in this study are unique between the F1, F2, and F3 generations.
The number of lncRNA was much higher than the number of sncRNA [small noncoding RNA, including microRNA]. The overlap between each generation was very low or nonexistent.
The F1 and the F2 generation control versus vinclozolin lineage sperm had negligible DHRs [differential histone retention sites]. This observation suggests that the direct vinclozolin exposure does not alter histone retention or trigger any changes. However, the F3 generation control versus vinclozolin lineage sperm DHRs increased considerably.
It appears that the phenomenon is more complex than just a direct exposure triggering the formation of epimutations that are then simply maintained in the subsequent generations.”
I asked the coauthors why a third of the cited references were self-referencing. The lead author replied:
“The field in epigenetic transgenerational inheritance is expanding, however it is still hard for us to find relevant studies in rodents or human that we can cite. Most of the time DNA methylation, ncRNA and histone modifications are investigated from a direct exposure and/or from a purely mechanistic angle (e.g. DNA methylation of specific genes).
In contrast, transgenerational phenotypes and toxicology by definition excludes direct exposure and must be transmitted through multiple generations (the F3 generation is the first transgenerational one). We are not looking at specific genes but using whole genome sequencing technologies which is a broader approach.
Besides, if you do a pubmed search with the keywords “epigenetics” and “transgenerational”, you will probably find that more than 50% of the studies have been done by Dr Michael K. Skinner. He is also one of the first researcher who started to work on the epigenetic transgenerational inheritance phenomenon 15 years ago. Not citing his previous work is challenging.
We hope to see other labs contributing to this particular field and we will be delighted to cite them. In the meantime, our only option is to reference our previous work.”
I replied:
“Thank you for your reply! It must be exasperating to see other researchers stop their studies short of the F3 generation for no apparent or disclosed reason.
Have you seen even one scientifically adequate human study of transgenerational epigenetic inheritance?”
https://academic.oup.com/eep/article/4/2/dvy010/4987173 “Alterations in sperm DNA methylation, non-coding RNA expression, and histone retention mediate vinclozolin-induced epigenetic transgenerational inheritance of disease”
This 2018 German review subject was detecting DNA modifications that are derivatives of the much-studied 5-methlycytosine:
“The discovery of modified nucleobases arising from 5-methylcytosine (5mC) through consecutive oxidation to give 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC), and 5-carboxylcytosine (5caC) has stimulated intense research efforts regarding the biological functions of these epigenetic marks.
Recent findings revealed that 5hmC and 5fC are stable DNA modifications in the genome, thus suggesting that oxidized 5mC derivatives may function as epigenetic marks in their own right, exhibiting regulatory purposes and participating in DNA replication, transcription, repair, and recombination.
The bisulfite-sequencing method (BS-Seq) has widely been used as the gold standard in determining the methylation status with single-base resolution in genomic DNA. The BS-Seq method, however, has some severe drawbacks, such as:
Harsh reaction conditions which might cause undesired DNA damage,
Requirements for relatively large amounts of input DNA,
Dependence on PCR, and resulting short sequence reads, as well as
Reduced sequence complexity due to deamination of all nonmethylated cytosines and
Accompanied challenges for primer hybridization.
Most importantly, however, with BS-Seq it is not possible to discriminate between 5mC and 5hmC..Furthermore, since 5fC and 5aC undergo deamination similar to unmodified cytosine, they are indistinguishable from C under bisulfite conditions.”