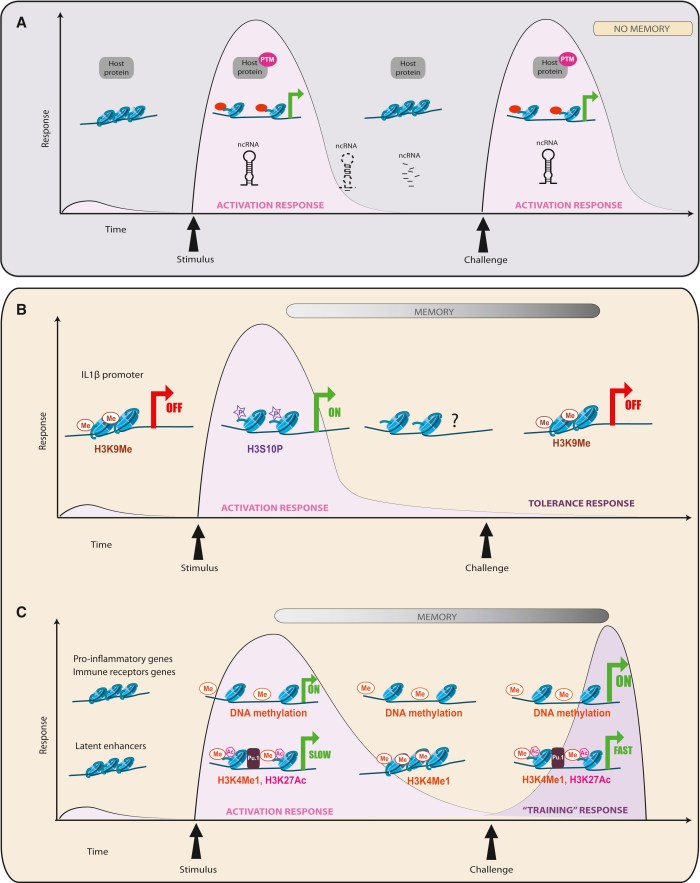
This 2016 Italian review subject was the interplay of genetic imprinting and sleep regulation:
“Sleep results from the synergism between at least two major processes: a homeostatic regulatory mechanism that depends on the accumulation of the sleep drive during wakefulness, and a circadian self-sustained mechanism that sets the time for sleeping and waking throughout the 24-hour daily cycle.
REM sleep apparently contravenes the restorative aspects of sleep; however, the function of this ‘paradoxical’ state remains unknown. Although REM sleep may serve important functions, a lack of REM sleep has no major consequences for survival in humans; however, severe detrimental effects have been observed in rats.
Opposite imprinting defects at chromosome 15q11–13 are responsible for opposite sleep phenotypes as well as opposite neurodevelopmental abnormalities, namely the Prader-Willi syndrome (PWS) and the Angelman syndrome (AS). Whilst the PWS is due to loss of paternal expression of alleles, the AS is due to loss of maternal expression.
Maternal additions or paternal deletions of alleles at chromosome 15q11–13 are characterized by temperature control abnormalities, excessive sleepiness, and specific sleep architecture changes, particularly REM sleep deficits. Conversely, paternal additions or maternal deletions at chromosome 15q11–13 are characterized by reductions in sleep and frequent and prolonged night wakings.
The ‘genomic imprinting hypothesis of sleep’ remains in its infancy, and several aspects require attention and further investigation.”
http://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1006004 “Genomic Imprinting: A New Epigenetic Perspective of Sleep Regulation”
A commenter to the review referenced a 2014 study Troubled sleep: night waking, breastfeeding, and parent–offspring conflict that received several reactions, including one by the same commenter. Here are a few quotes from the study author’s consolidated response:
“‘Troubled sleep’ had two major purposes. The first was to draw attention to the oppositely perturbed sleep of infants with PWS and AS and explore its evolutionary implications. The involvement of imprinted genes suggests that infant sleep has been subject to antagonistic selection on genes of maternal and paternal origin with genes of maternal origin favoring less disrupted sleep.
My second major purpose was a critique of the idea that children would be happier, healthier and better-adjusted if we could only return to natural methods of child care. This way of thinking is often accompanied by a belief that modern practices put children at risk of irrevocable harm.
The truth of such claims is ultimately an empirical question, but the claims are sometimes presented as if they had the imprimatur of evolutionary biology. This appeal to scientific authority often seems to misrepresent what evolutionary theory predicts: that which evolves is not necessarily that which is healthy.
Why should pregnancy not be more efficient and more robust than other physiological systems, rather than less? Crucial checks, balances and feedback controls are lacking in the shared physiology of the maternal–fetal unit.
Infant sleep may similarly lack the exquisite organization of systems without evolutionary conflict. Postnatal development, like prenatal development, is subject to difficulties of evolutionarily credible communication between mothers and offspring.”
The author addressed comments related to attachment theory:
“Infants are classified as having insecure-resistant attachment if they maintain close proximity to their mother after a brief separation while expressing negative emotions and exhibiting contradictory behaviors that seem to both encourage and resist interaction. By contrast, infants are classified as having insecure-avoidant attachment if they do not express negative emotion and avoid contact with their mother after reunion.
Insecure-avoidant and insecure-resistant behaviors might be considered antithetic accommodations of infants to less responsive mothers; the former associated with reduced demands on maternal attention, the latter with increased demands. A parallel pattern is seen in effects on maternal sleep. Insecure-avoidant infants wake their mothers less frequently, and insecure-resistant infants more frequently, than securely attached infants.
Parent–child interactions are transformed once children can speak. Infants with more fragmented sleep at 6 months had less language at 18 and 30 months.
Infants with AS have unconsolidated sleep and never learn to speak. The absence of language in the absence of expression of one or more MEGs [maternally expressed imprinted genes] is compatible with a hypothesis in which earlier development of language reduces infant demands on mothers.”
Regarding cultural differences:
“China, Taiwan and Hong Kong have both high rates of bed-sharing and high rates of problematic sleep compared with western countries. Within this grouping, however, more children sleep in their own room but parents report fewer sleep problems in Hong Kong than in either China or Taiwan.
Clearly, cultural differences are significant, and the causes of this variation should be investigated, but the differences cannot be summarized simply as ‘west is worst’.
The fitness [genetic rather than physical fitness] gain to mothers of an extra child and the benefits for infants of longer IBIs [interbirth intervals] are substantial. These selective forces are unlikely to be orders of magnitude weaker than the advantages of lactase persistence, yet the selective forces associated with dairying have been sufficient to result in adaptive genetic differentiation among populations.
The possibility of gene–culture coevolution should not be discounted for behaviors associated with infant-care practices.”
Regarding a mismatch between modern and ancestral environments:
“I remain skeptical of a tendency to ascribe most modern woes to incongruence between our evolved nature and western cultural practices. We did not evolve to be happy or healthy but to leave genetic descendants, and an undue emphasis on mismatch risks conflating health and fitness.
McKenna [a commenter] writes ‘It isn’t really nice nor maybe even possible to fool mother nature.’ Here I disagree. Our genetic adaptations often try to fool us into doing things that enhance fitness at costs to our happiness.
Our genes do not care about us and we should have no compunction about fooling them to deliver benefits without serving their ends. Contraception, to take one obvious example, allows those who choose childlessness to enjoy the pleasures of sexual activity without the fitness-enhancing risk of conception.
Night waking evolved in environments in which there were strong fitness costs from short IBIs and in which parents lacked artificial means of birth-spacing. If night waking evolved because it prolonged IBIs, then it may no longer serve the ends for which it evolved.
Nevertheless, optimal infant development might continue to depend on frequent night feeds as part of our ingrained evolutionary heritage.
It could also be argued that when night waking is not reinforced by feeding, and infants sleep through the night, then conflict within their genomes subsides. Infants would then gain the benefit of unfragmented sleep without the pleiotropic costs of intragenomic conflict. Plausible arguments could be presented for either hypothesis and a choice between them must await discriminating evidence.”
Commenters on the 2014 study also said:
“[Crespi] The profound implications of Haig’s insights into the roles of evolutionary conflicts in fetal, infant and maternal health are matched only by the remarkable absence of understanding, appreciation or application of such evolutionary principles among the research and clinical medical communities, or the general public.
[Wilkins] A mutation may be selected for its effect on the trait that is the basis of the conflict, but that mutation also likely affects other traits. In general, we expect that these pleiotropic effects to be deleterious: conflict over one trait can actually drive other traits to be less adapted. Natural selection does not necessarily guarantee positive health outcomes.
[McNamara] Assuming that AS/REM is differentially influenced by genes of paternal origin then both REM properties and REM-associated awakenings can be better explained by mechanisms of genomic conflict than by traditional claims that REM functions as an anti-predator ‘sentinel’ for the sleeping organism.
[Hinde] Given this context of simultaneous coordination and conflict between mother and infant, distinguishing honest signals of infant need from self-interested, care-extracting signals poses a challenge.“