This 2016 Oregon review’s concept was the inadequacy of drug-based therapies, explored with the specific subject of epilepsy:
“Currently used antiepileptic drugs:
- [aren’t] effective in over 30% of patients
- [don’t] affect the comorbidities of epilepsy
- [don’t] prevent the development and progression of epilepsy (epileptogenesis).
Prevention of epilepsy and its progression [requires] novel conceptual advances.”
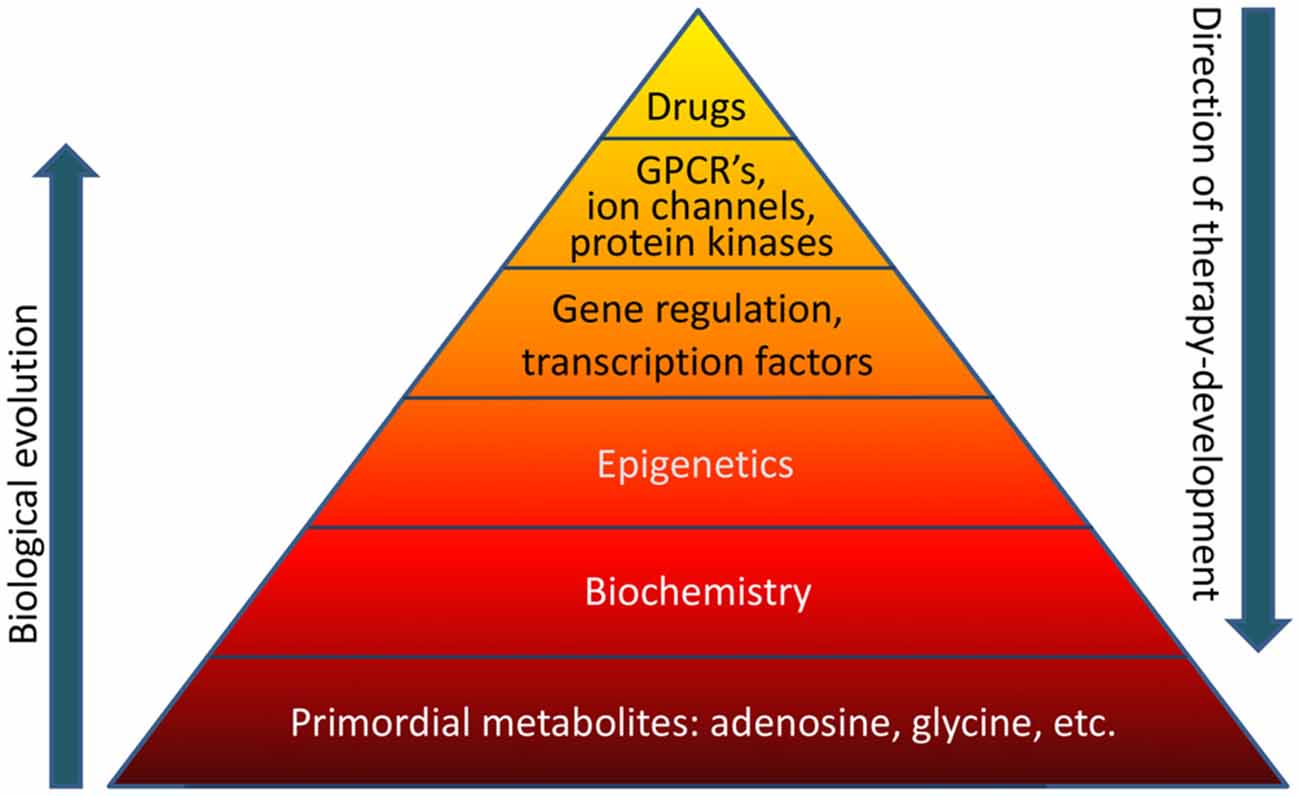
The overall concept that current drug-based therapies poorly address evolutionary biological realities was illustrated by a pyramid, with the comment that:
“If the basis of the pyramid depicted in Figure 1 is overlooked, it becomes obvious that a traditional pharmacological top-down treatment approach has limitations.”
I would have liked the reviewer to further address the “therapeutic reconstruction of the epigenome” point he made in the Abstract:
“New findings based on biochemical manipulation of the DNA methylome suggest that:
- Epigenetic mechanisms play a functional role in epileptogenesis; and
- Therapeutic reconstruction of the epigenome is an effective antiepileptogenic therapy.”
As it was, the reviewer lapsed into the prevalent belief that the causes of and cures for human diseases will always be found on the molecular level – for example, the base of the above pyramid – and never in human experiences. This preconception leads to discounting human elements – notably absent in the above pyramid – that generate epigenetic changes.
A consequence of ignoring experiential causes of diseases is that the potential of experiential therapies to effect “therapeutic reconstruction of the epigenome” isn’t investigated.
http://journal.frontiersin.org/article/10.3389/fnmol.2016.00026/full “The Biochemistry and Epigenetics of Epilepsy: Focus on Adenosine and Glycine”

