People will forgive you for being wrong, but they will never forgive you for being right – especially if events prove you right while proving them wrong. Thomas Sowell
1. I’ve continued daily practices from Year Five to experience another year without being sick (if I don’t count getting MSG poisoning from Chinese food.) I consequently scheduled a doctor visit next week to get a sumatriptan prescription refilled.
– In that post’s comments, Ole Bisgaard Pedersen asked if I took NAD+ supplements such as NAM, NMN or NR – forms of vitamin B3 that are precursors to NAD+. I didn’t note that last year I started taking Now brand Flush-free niacin 500 mg mid-morning.
Nicotinamide riboside has the most human evidence, including a Tru Niagen clinical trial that showed improvements in peripheral artery disease. It’s too expensive for long-term use, though.
Even if I could afford it, there isn’t a magic bullet for fixing vascular system dysfunction. PAD is just one symptom of a cardiovascular system that needs to be overhauled then maintained at a healthy level. There is no clinical trial that has a logical therapeutic end point to stop treatments where a person could say, “I’ve done enough for my vascular system, my physical and cognitive functions won’t backslide.”
– 2-3 years ago, I changed from microwaving broccoli sprouts in a plastic bag to microwaving them in a small bowl with a small plate covering it to keep them from popcorning out of the bowl. I use a 1000W microwave oven on 80% power for ten seconds.
3. The two vitamin C macaque studies I’ve recently curated both ran for a human equivalent of ten years. I was encouraged that both found Nrf2 activation to be part of their causal beneficial evidence, since vitamin C wasn’t on my radar as a Nrf2 activator.
I expect that in four years I’ll write a Year Ten post on eating microwaved broccoli sprouts. I haven’t seen human evidence for broccoli extracts that bypass small intestine absorption and metabolism per Glucosinolate and isothiocyanate human interventions, or enteric capsules, or nanoformulations as suitable substitutes. Maybe studies on broccoli sprout powder or a Nrf2 activator that tops sulforaphane will be published before then, who knows.
I switch things around pretty often, but I haven’t said much about diet and supplement changes since this time last year. Here’s what I’ve done in terms of changes that I’ve since abandoned or reduced, followed by additions or increases that I’ve kept.
Abandoned and reduced items
1. I stopped using Avena sativa oats to grow 3-day-old oats sprouts. I again ran into the same situation where I got < 10% yield.
The first time this happened in 2023, I related to the Montana farmer that degraded seed vitality was probably caused by the way that Amazon handled their oat products. I’m the customer, though, and I won’t make it my problem if the vendor can’t meet expectations.
I switched to sprouting Avena nuda oats based on Sprouting hulless oats. I’ll note that this Illinois farmer doesn’t let Amazon handle their organic Avena nuda oats, and they add on post office shipping costs. They don’t recommend sprouting, probably because of liability, although I’ve had a 91% germination rate over three days. I might have ordered Avena sativa oats directly from the Montana farmer bypassing Amazon if they were also organic.
2. I stopped taking alpha ketoglutarate. In my view, increasing tricarboxylic acid (TCA) cycle intermediate metabolites such as alpha ketoglutarate and CoQ10 should not be the primary way to improve mitochondrial electron transport chain function.
Instead of biochemical considerations, focus on photon modulation, which precedes biochemical reactions. Which means mitochondrial studies should be controlled for light exposures, and very few of them do that, although it’s the way nature works.
This past winter I increased indoor non-LED light exposure within a circadian rhythm framework. I’ll switch back to walking the beach at sunrise from being out in mid-day sun after it gets a little bit warmer.
3. I’ve taken creatine on and off during the past year. There’s a bit of literature on its use for improving methyltransferase system components like homocysteine.
Stopping creatine fits one of the overall patterns that studies demonstrate – people who are initially deficient in the studied item get a benefit, while people who are initially sufficient don’t benefit from treatment. I’ve always tested mid-range for homocysteine, which is desirable.
4. I had some cocoa powder lying around for a year or two, and I used it this past winter to improve the taste of coffee I bought on sale. Cocoa flavanols are supposed to improve various health measures. But I haven’t been provided access to the most recent human studies, so I won’t repeat their results without reading their details.
For me, though, the dryness of a chewed pecan bolus creates a swallowing problem that walnuts don’t have. YMMV.
6. I stopped taking 2 g magnesium L-threonate. I’ve always tested high for magnesium without using a specific supplement.
7. I reduced D3 by 25 mcg to a daily 2400 IU. Winter is over.
New and increased items
1. I curated five 2025 ergothioneine studies in Human studies of ergothioneine after stopping mushroom intake via AGE-less chicken soup. I wasn’t thrilled that none of them investigated long-term effects of persistent plasma ergothioneine levels.
This year I decided to start taking the higher 25 mg dose of the first study once a week. That should produce some benefits at a lower ergothioneine blood level than daily doses produce. I’ll check periodically for 2026 research.
2. The only paper I’ve curated on deuterium (heavy hydrogen) is Taurine and mitochondrial health. I started using Icelandic glacier water to make coffee and tea, and for just drinking.
It isn’t advertised as deuterium-depleted water, and it isn’t manufactured as such. But I think any glacier water contains less deuterium than local water. I use local filtered water for sprouting and cooking.
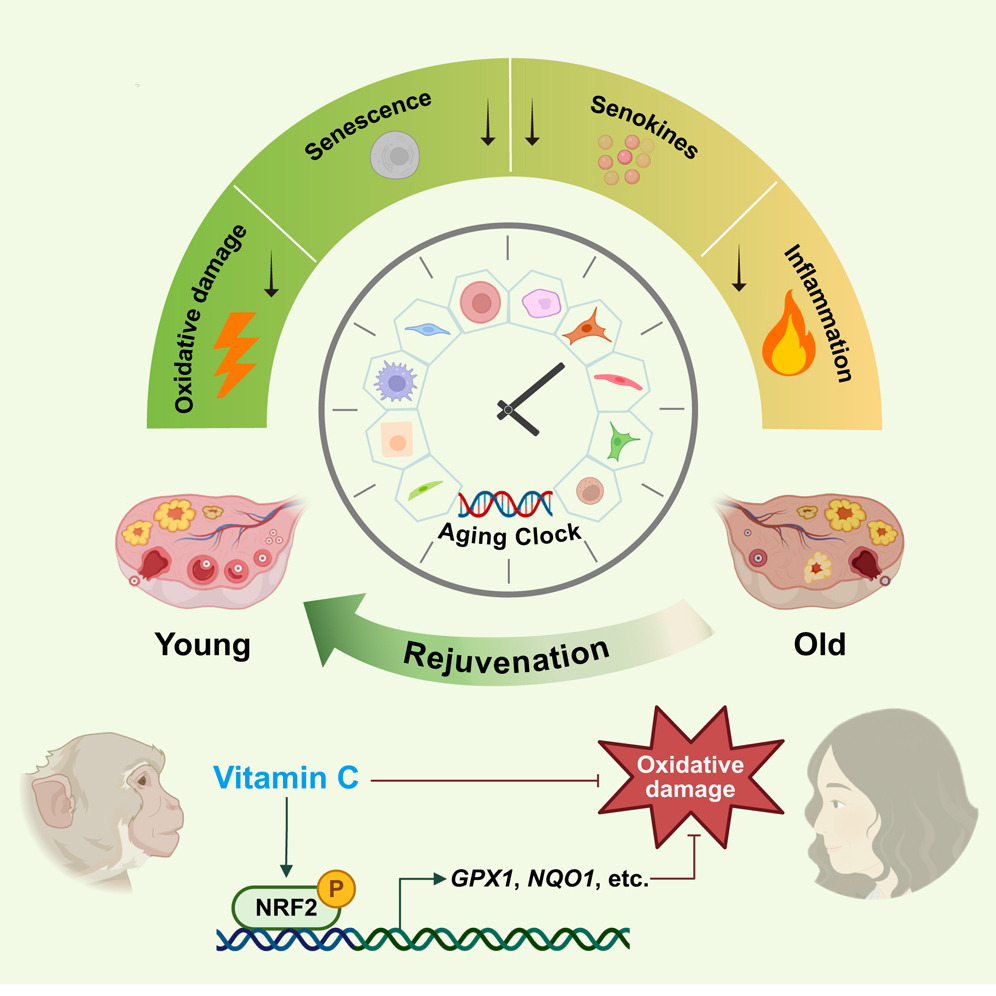
3. Per The return of the free radical theory of aging I started taking extra vitamin C separately from other supplements in the form of Now brand liposomal 1 gram twice daily this past winter. That study found vitamin C to be an anti-aging compound for primates.
“VC slowed aging in various ovarian cell types. Moreover, VC protected human ovarian endothelial and stromal cells (SCs) from aging partially via NRF2 activation. This study establishes a proof-of-concept for delaying primate ovarian aging with a single compound, and provides important insights into preventing and treating degenerative diseases related to ovarian aging.”
4. I restarted taking inulin last year, about 3 grams (a heaping teaspoon) daily after posting Inulin vs. FOS. My 2.5 year-old grandchild takes a level teaspoon daily, as inulin’s beneficial effects aren’t just for old people.
5. I started taking 12 mg astaxanthin twice in the morning. I use Nrf2 activators in the morning because Nrf2 is especially involved in the circadian cycle, as noted in papers such as Broccoli sprouts activate the AMPK pathway, Part 4.
6. I increased daily raw egg consumption from 3 eggs a day to 3 eggs twice daily.
7. This year, Ovega 3 algae oil DHA 420 mg/EPA 140 mg became no longer available AFAIK. I substituted Vegan Omega 3 algae oil DHA 300 mg/EPA 150 mg in the morning and Sports Research Omega 3 fish oil DHA 310 mg/EPA 690 mg in the afternoon.
8. I picked up this Korean seaweed in a 10-pack at Costco. The label doesn’t say what its iodine content is. I eat it as a snack whenever I get a salt craving, maybe once a week.
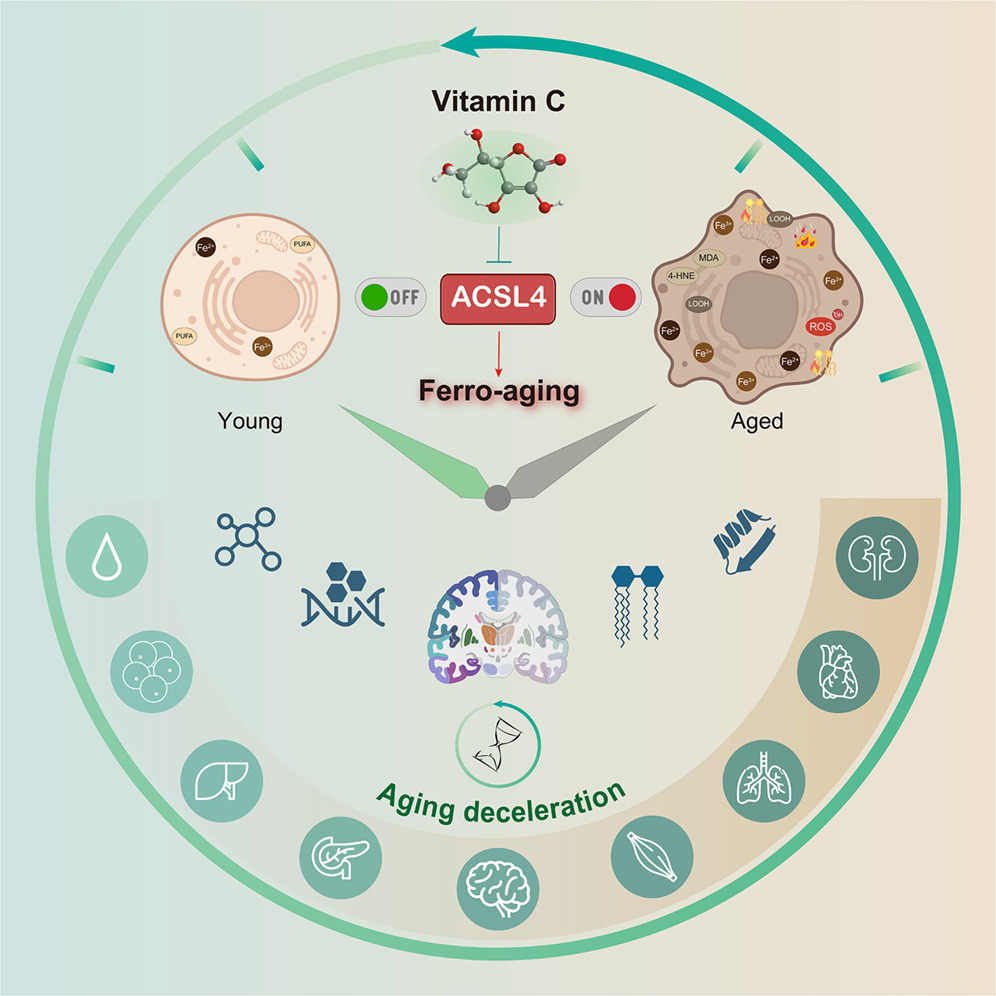
A 2026 primate study investigated effects of vitamin C:
“Here, we define a conserved iron-lipid axis driving primate aging, termed ‘ferro-aging.’ Multi-tissue profiling in humans and non-human primates reveals age-progressive iron accumulation, fueling chronic lipid peroxidation orchestrated by acyl-coenzyme A (CoA) synthetase long-chain family member 4 (ACSL 4). Distinct from acute ferroptosis, this ACSL4-mediated process promotes cellular senescence and systemic functional decline.
We identify vitamin C (VC) as a direct inhibitor of ACSL4. Long-term VC administration in aged monkeys for over 40 months potently reduces ferro-aging signatures across tissues, attenuates multi-organ pathology, and improves neurological and metabolic functions. Multi-omic aging clocks indicate the VC-mediated reversal of biological age.
Despite decades of interest in oxidative stress, largely sparked by the free radical theory of aging, efforts to modulate it broadly with antioxidants have yielded inconsistent or neutral outcomes, highlighting the theory’s limitations and underscoring the need to identify more specific, upstream drivers. A critical challenge remains: determining whether the iron-lipid axis constitutes a core upstream driver of aging in primates and, if so, whether it is therapeutically targetable.
In this study, we bridge these gaps. We define an iron-triggered, ACSL4-governed, lipid peroxidation-driven program that escalates with age across diverse cell types and multiple organs in non-human primates.
VC treatment dose-dependently increased Nrf2 phosphorylation and activation. VC orchestrates a dual-defense strategy against ferro-aging: it directly suppresses the pro-aging lipid peroxidation driver ACSL4, while in parallel, it bolsters the cell’s intrinsic antioxidant capacity via Nrf2 pathway activation.
Middle-aged cynomolgus monkeys (12–16 years old, approximating human 40–50 years) received daily oral VC (30 mg/kg group) or a control treatment for 40 months under standardized conditions.
Structural MRI analysis demonstrated that VC intervention counteracted age-related brain atrophy. Using general linear mixed models, we found that VC restored cortical surface area in the frontal lobes of aged monkeys. Regional analysis identified enlargement in four regions of the orbital frontal cortex, an area critical for adaptivebehavior.
Diffusion MRI-based connectomics revealed that, compared with young animals, aged monkeys exhibited reduced structural connectivity in 18 brain regions. VC treatment restored connectivity in 9 of these regions, which were predominantly located in the posterior parietal cortex, a hub for spatial awareness and decision-making.
VC exerted robust neuroprotective effects. It attenuated heterochromatin loss (increased H3K9me3) in the prefrontal cortex and hippocampus and reduced abnormal protein aggregates, including cytosolic aggresomes and Aβ. Additionally, VC lowered the abundance of activated microglia and astrocytes and suppressed expression of the innate immune sensor cGAS in the hippocampus.
VC supplementation reduced the estimated biological age across multiple organs. At the epigenetic level, VC lowered DNA methylation age in several tissues, including brain, brown adipose tissue, muscle, skin, aorta, and kidney. In the hippocampus, the most substantial reductions in biological age occurred in microglia, oligodendroglia, and oligodendrocyte precursor cells. In the pancreas, alpha cells, beta cells, and ductal cells showed the greatest rejuvenation.
In summary, chronic VC supplementation inhibits the ferro-aging pathway, reduces multidimensional biological age across primate organs, and ameliorates a spectrum of functional declines in nervous and metabolic systems. Our work establishes ACSL4 inhibition as a promising and translationally relevant therapeutic strategy for mitigating aging-related decline.
A long-term, 40-month intervention study in aged non-human primates is a highly translational model given their shared inability with humans to synthesize VC endogenously. The finding that a single, safe nutrient can reverse multidimensional aging clocks in a primate has profound implications for translational longevity medicine.“
“For humans (who, like macaques, cannot synthesize vitamin C), the Recommended Dietary Allowance (RDA) is 75–90 mg/day for adults (~1–1.5 mg/kg for a 60–70 kg person) to prevent deficiency. Upper safe intake levels are much higher: up to 2,000 mg/day (Tolerable Upper Intake Level) is considered safe for most adults, with no established adverse effects at that level from food/supplements.
Treated monkeys represent advanced aging stages (likely equivalent to human 50s–70s+ based on ‘aged’ designation and long-term intervention effects), extending the prior 12–16-year monkey range (human ~35–55) to broader anti-aging applications. While human trials are needed, the primate evidence (long-duration, systemic benefits) strengthens the case for high-dose, sustained vitamin C as a strategy against ferro-aging in humans. It elevates vitamin C from a nutrient to a targeted anti-aging compound in primates.”
Coincidentally, I started taking extra vitamin C separately from other supplements in the form of liposomal 1 gram twice daily this past winter. Can’t say that it had any effects on my intended target, avoiding sniffles and sneezing, as allergy season kicked off in early February. With this study’s findings, I’ll continue.
A 2026 rodent study investigated sulforaphane’s ability to affect ALS-like symptoms:
“The objective of this study was to evaluate neuroprotective efficacy and safety of sulforaphane (SUFP) in a methylmercury (MMHg⁺)-induced preclinical rat model of amyotrophic lateral sclerosis (ALS). ALS is characterized by progressive motor neuron degeneration and muscle wasting, leading to impairments in gait, swallowing, salivation, and routine motor activities.
64 animals were classified into eight groups: 1st: normal control, 2nd: vehicle control; 3rd: SUFP perse (4 mg/kg, i.p.), 4th: MMHg + (5 mg/kg, p.o.), 5th: MMHg + 5 + SUFP (2 mg/kg, i.p.), 6th: MMHg+ 5 + SUFP (4 mg/kg, i.p.), 7th: MMHg+ 5 + omaveloxolone (OVX) (30 mg/kg, i.p.), and 8th: MMHg + 5 + dimethyl fumarate (DIMT) (50 mg/kg, i.p.). Neurotoxin MMHg + was orally administered at 5 mg/kg for the first 21 days. For the next 22 days, SUFP, OVX, and DIMT were administered intraperitoneally (i.p.).
SUFP modulates neurotransmitter levels such as acetylcholine (A), dopamine (B), GABA (C), glutamate (D), and serotonin (E).
SUFP4 exerted broad neuroprotective effects in ALS pathology by restoring antioxidant proteins (Nrf2, HO-1, SIRT1), suppressing apoptotic (Bax, caspase-3, Bcl-2) and inflammatory markers (TNF-α, IL-1β), and enhancing the anti-inflammatory cytokine IL-10. It also downregulated stress-related signaling pathways (PI3K/Akt, p75NTRECD, MAPKs) associated with neurodegeneration. These molecular effects translated into meaningful functional recovery, as evidenced by improvements in grip strength, locomotor performance, spatial memory, and depressive-like behavior.
Histopathological evaluation demonstrated attenuation of demyelination and preservation of neuronal architecture including the cerebral cortex, hippocampus, striatum, midbrain, and cerebellum. Beyond central neuroprotection, SUFP exerted systemic benefits by normalizing hepatic enzymes, improving skeletal muscle integrity, restoring redox balance, stabilizing neurofilament and myelin-associated proteins, and correcting hematological alterations.
Despite limitations related to study duration and animal sex, this work strongly positions SUFP as a promising, multi-target therapeutic candidate for ALS with both neural and systemic protective efficacy.”
https://link.springer.com/article/10.1007/s12035-026-05683-5 “Sulforaphane-Mediated Multitarget Therapeutic Effects in Methylmercury-Induced ALS-Like Pathology: Comparative Analysis and Multifaceted Approach to Neuroprotection and Systemic Recovery” (not freely available) Thanks to Dr. Sidharth Mehan for providing a copy.
Unlike A Nrf2 treatment for ALS?, this study didn’t present evidence that its treatment compound was effective for preventing ALS. For one thing, currently-known disease factors involving heat shock proteins and associated genes, some of which are Nrf2 targets, weren’t investigated.
Two Nrf2 activators were used in both studies as comparators of Nrf2 activation effects. Neither omaveloxolone nor dimethyl fumarate are ALS causal treatments, though, and have undesirable side effects.
A human equivalent of this study’s higher sulforaphane dose is ((4 mg x .162) x 70 kg) = 45 mg. 45 mg of sulforaphane might be too much to consistently take at one time because of unpalatability. But I documented taking an estimated 52 mg for a year during 2020-2021 by eating microwaved 3-day-old broccoli sprouts twice a day.
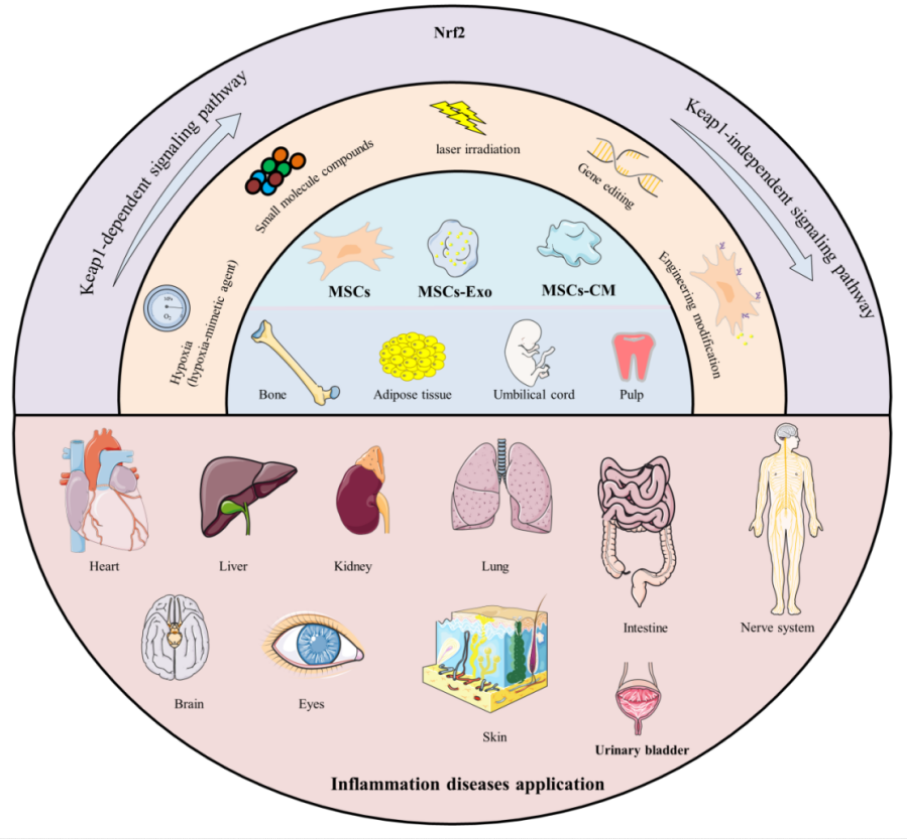
A 2026 review subject was mechanisms and therapeutic potential for Nrf2 activators in combination with mesenchymal stem cells:
“Mesenchymal stromal/stem cells (MSCs) are multipotent stem cells that can be isolated from various tissues – such as bone marrow (BM), umbilical cord (UC), adipose tissue (AD), dental pulp (DP), hair follicle (HF), and placenta – and differentiated into multiple lineages under appropriate conditions. Their functional repertoire includes immunomodulation, homing, and differentiation, which collectively help establish a balanced inflammatory and regenerative niche within damaged tissues during severe inflammation. MSCs-derived extracellular vesicles (MSCs-EVs) and conditioned medium (MSCs-CM) play remarkable roles, exhibiting potent anti-inflammatory and antioxidant properties that offer novel therapeutic alternatives for inflammatory diseases.
Therapeutic capacity of MSCs in inflammatory conditions is increasingly attributed to their potent paracrine activity rather than solely to their differentiation potential. A key mechanism underlying this paracrine effect is activation of the Nrf2 antioxidant pathway.
MSCs and their secreted products including exosomes (Exos), EVs, and CM, activate Nrf2 through multi-dimensional/target mechanisms, thereby enhancing cellular antioxidant defenses, modulating immune responses, and promoting tissue repair. It is noteworthy that therapeutic efficacy of MSCs and their derivatives can be enhanced through external modulation, including pretreatment with natural compounds.
Preconditioning refers to brief treatment of MSCs or their derivatives with physical, chemical, or biological factors prior to application, aiming to enhance their ability to counteract oxidative stress and improve their therapeutic efficacy. Flavonoids precondition and prime MSCs via the direct Keap1-Nrf2 pathway or indirect PI3K-Akt pathway, which enhances cellular resilience to adverse conditions by reducing apoptosis and promoting survival. Primed MSCs, in turn, remodel the microenvironment through an altered secretory profile, releasing bioactive factors that create more favorable conditions for their own persistence.
The core logic of these strategies lies in simulating or inducing adaptive stress, such as employing specific chemical molecules or drug stimuli, or utilizing physical / microenvironmental preconditioning to mimic specific physical conditions of the in vivo injury environment. The most straightforward strategy is overexpression of Nrf2 or its key downstream effector molecules.
The majority of existing studies remain at the level of observing correlations with Nrf2 upregulation, and there is still a lack of precise causal validation regarding key upstream signals – such as specific cytokines, miRNAs, or proteins – through which MSCs or derivatives initiate Nrf2 activation. Mechanistic insights are predominantly derived from in vivo or rodent (mouse/rat) model experiments, with a notable absence of clinical validation, insufficient long-term safety and pharmacokinetic data, and a lack of standardization in administration routes and dosages, all of which hinder clinical translation.
The essential role of the Nrf2 pathway has not been rigorously confirmed, as most studies have not employed reverse genetic validation using Nrf2-knockout animals or specific inhibitors. Consequently, it remains unclear whether therapeutic effects are necessarily and exclusively dependent on Nrf2, and potential synergistic contributions from other pathways may have been overlooked.
Most natural flavonoids face challenges such as low oral bioavailability, rapid metabolism, and poor targeting. Numerous challenges remain to be addressed in order to translate these promising preclinical findings into clinical practice. Future research should focus on the following aspects:
Elucidating precise upstream molecular mechanisms by which MSCs activate Nrf2;
Employing more clinically relevant chronic disorder models;
Systematically evaluating long-term safety, optimal delivery strategies (including dosage and route of administration), and immunogenicity of MSCs-based therapies;
Validating selection criteria (optimal source), quality control, batch-to-batch consistency of MSCs, and addressing regulatory and ethical barriers to clinical translation; and
Integrating molecular docking, ADMET (Absorption, Distribution, Metabolism, Excretion, Toxicity) prediction, and in vitro and in vivo validation to further elucidate regulatory effects of flavonoids and enhance understanding of their mechanisms of action.”
This paper was overly long at 127 pages, so I focused on the later sections. None of these treatments are currently ready for clinical trials.
I also didn’t mention specific flavonoids as Nrf2 activators. It’s beyond a reviewer’s task to rank Nrf2 activators, and a study’s researchers seldom address why they used a poorly-activating flavonoid instead of a higher-ranked natural plant compound such as sulforaphane.
A 2026 paper provided details of a 2020-2023 human trial of broccoli sprouts:
“In a 42-month randomized, double-blind, placebo-controlled trial, 26 participants aged 63–90 years with memory impairment were randomly assigned to receive either 30 mg/day of glucoraphanin (GLR) or placebo. The primary outcome was the change in Memory Performance Index (MPI) scores from the mild cognitive impairment (MCI) screen. This study evaluated the long-term efficacy of GLR supplementation on cognitive function in older adults at an elevated risk for Alzheimer’s disease (AD), including those with MCI.
Participants were instructed to take three capsules of either the GLR or placebo supplements daily for 42 months. The GLR supplement contained 30 mg of GLR purified from broccoli sprouts, along with 120 mg of mustard powder per three capsules. Mustard powder was included as a source of exogenous active myrosinase to enhance the enzymatic conversion of GLR to sulforaphane. The placebo supplement contained 0 mg of GLR.
No significant group difference was observed in the initial 6 months. A marginal difference in favor of GLR appeared in the later phase (30 and 42 months), including the 42-month endpoint.
The GLR group demonstrated superior performance on immediate recall and delayed free recall tests. MCI participants showed a greater MPI improvement with GLR.
Long-term GLR supplementation may help preserve cognitive function in individuals at elevated risk for AD, particularly those with MCI. Larger trials are warranted to confirm efficacy and clarify underlying mechanisms.”
This study was funded by the supplement manufacturer. There was no explanation of what the supplement’s “purified from broccoli sprouts” entails. Also, I didn’t mention results of voluntary group exercise because there was a long gap in the participants’ data due to government response to covid.
For comparison of this study’s 30 mg glucoraphanin dose, Our model clinical trial for Changing to a youthful phenotype with broccoli sprouts provided 30 grams of fresh broccoli sprouts that contained an estimated 51 mg of glucoraphanin for ten weeks. That study’s corresponding coauthor said of their 30 gram broccoli sprouts dose in Understanding a clinical trial’s broccoli sprout amount that “When we carried out tests with consumers, previous to the bioavailability studies, higher amounts per day were not easy to consume and to get eaten by participants.” There was no rationale provided for this study’s 30 mg dose other than citing two previous human studies that also used a 30 mg glucoraphanin dose.
For comparison of this study’s 120 mg mustard seed powder dose, my daily cruciferous food intake since five and a half years ago includes sprouted yellow mustard seeds started with 3.5 grams of seeds, along with sprouted broccoli and red cabbage started with 3.6 grams of each vegetable’s seeds, all sprouted for three days. I haven’t seen studies that show sprouting has effects on myrosinase enzyme activity.
This study cited Does sulforaphane reach the colon? which used 2% mustard seed powder to create sulforaphane from glucoraphanin. This study’s 120 mg mustard seed powder / 30 mg glucoraphanin is a lot more than 2%. My daily sprout intake started from 3.5 g mustard seeds / (3.6 g broccoli seeds +3.6 g red cabbage seeds) is also a lot more than 2%.
I’ve changed some items along the way, switching supplier from True Leaf to Johnny’s for organic broccoli seeds, and from non-organic Red Acre red cabbage seeds to True Leaf organic red cabbage seeds. I recently had to find another supplier of organic yellow mustard seeds when Naturevibe stopped carrying that product. I tried Food to Live, but their yellow mustard seeds when sprouted aren’t mild. I’ll next try Frontier Co-op to see if those are mild as advertised.
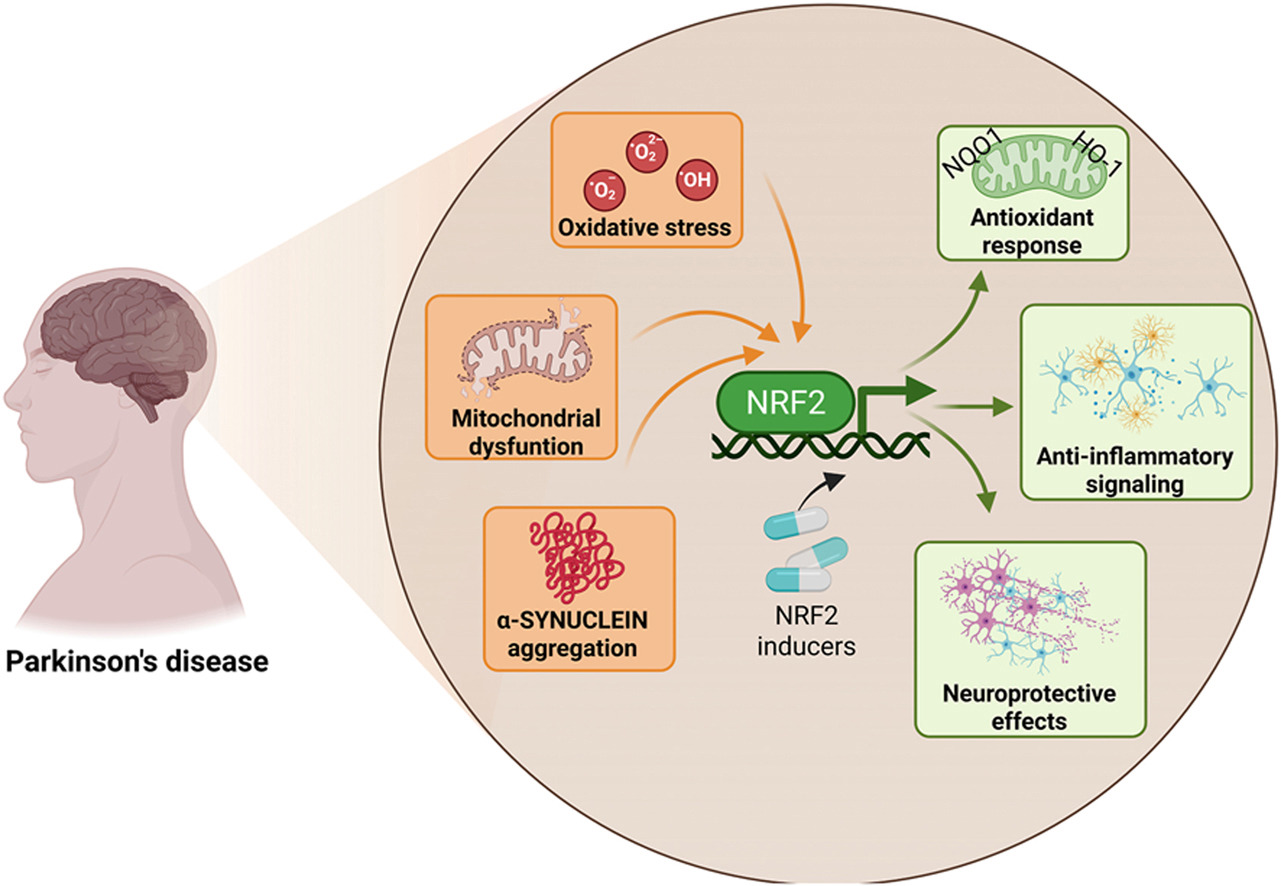
Starting this blog’s twelfth year by curating a poorly-done 2026 review of Nrf2 and its capability to change a person’s development of Parkinson’s disease. I’ll emphasize precedent conditions that if not effectively dealt with in youth, can’t prevent PD from occurring at some later life stage.
“This review explicitly examines how age-associated decline in NRF2 responsiveness intersects with redox imbalance, mitochondrial dysfunction, proteostatic failure, and neuroinflammation, core mechanisms shared between aging and PD. PD unfolds through a complex interplay of cellular stress and immune responses. Oxidative stress, mitochondrial dysfunction, and chronic neuroinflammation converge to damage dopaminergic neurons, with microglia playing a central role in amplifying this injury.
NRF2 emerges as a key regulator of antioxidant defenses, inflammatory balance, and mitochondrial protection, offering a promising target for clinical intervention. NRF2 activity favors the anti-inflammatory microglial over the pro-inflammatory phenotype. Decline in NRF2 inducibility with age impairs microglial clearance, promotes neuroinflammation, and reduces antioxidant defenses, while NRF2 activation restores protective functions and offers a promising therapeutic target.
Strategies aimed at restoring or enhancing NRF2 activity hold significant promise as disease-modifying interventions, not only to slow PD progression but also to promote resilience against the broader spectrum of age-associated neurodegenerative and inflammatory conditions.”
This review only gave lip service to PD progression outside of the brain, as if the importance of prodromal factors to a person’s neurodegeneration such as dysfunction in gut, eyes, skin, and olfactory systems can be minimized. But failure to recognize early what will doom a person to be unable to recover health in later decades is disingenuous. Since these reviewers omitted early interventions into PD prodromal factors, the best they came up with was interventions to “slow PD progression.”
Maybe these reviewers felt it would be outside the scope of this review to discuss early non-brain PD factors for more than one sentence? However, while PD is defined by striatal brain neurons, Nrf2 activity is much less in brain and central nervous system neurons than elsewhere in the body per Nrf2 Week #2: Neurons.
I disagree with these reviewers’ self-imposed emphasis on aging. Repeating ‘age-associated’ numerous times seemed as if they wanted to influence the reader into thinking age in and of itself was a cause for PD, rather than an imputed mathematical correlation. Their emphasis led to dumb mentions such as senolytics for no apparent reason than senescence is a ‘hallmark of aging’, and to meaningless ‘diseasome of aging’ characterizations, and to ignoring the existence of early non-age-associated PD diagnoses in 20- and 30-year-olds.
Whatever it takes to get published, I’d guess. Or maybe it’s that the number of omissions and useless points a review paper makes increases with the number of reviewers and their sponsors’ agendas.
For example, why was it permissible to dedicate lip service to ‘exposome’ factors like microplastics, environmental pollution, and viruses, but it’s still not permitted in 2026 to discuss research into the impacts on vascular disease and neurodegeneration of lipid nanoparticles and DNA contamination in what a large number of humans were exposed to by injected pharmaceuticals starting in late 2020? Not to mention two studies published in 2024 of over 2.5 million people whose incidences of neurologic issues, mild cognitive impairment, and Alzheimer’s disease rapidly increased after ‘vaccination’?
I’ve mentioned in this blog many times how it’s every human’s choice whether or not we take responsibility for our own one precious life. I suggest, if it’s not too late, do that for your children’s lives, too.
Here are five 2025 human ergothioneine studies, starting with a clinical trial of healthy older adults:
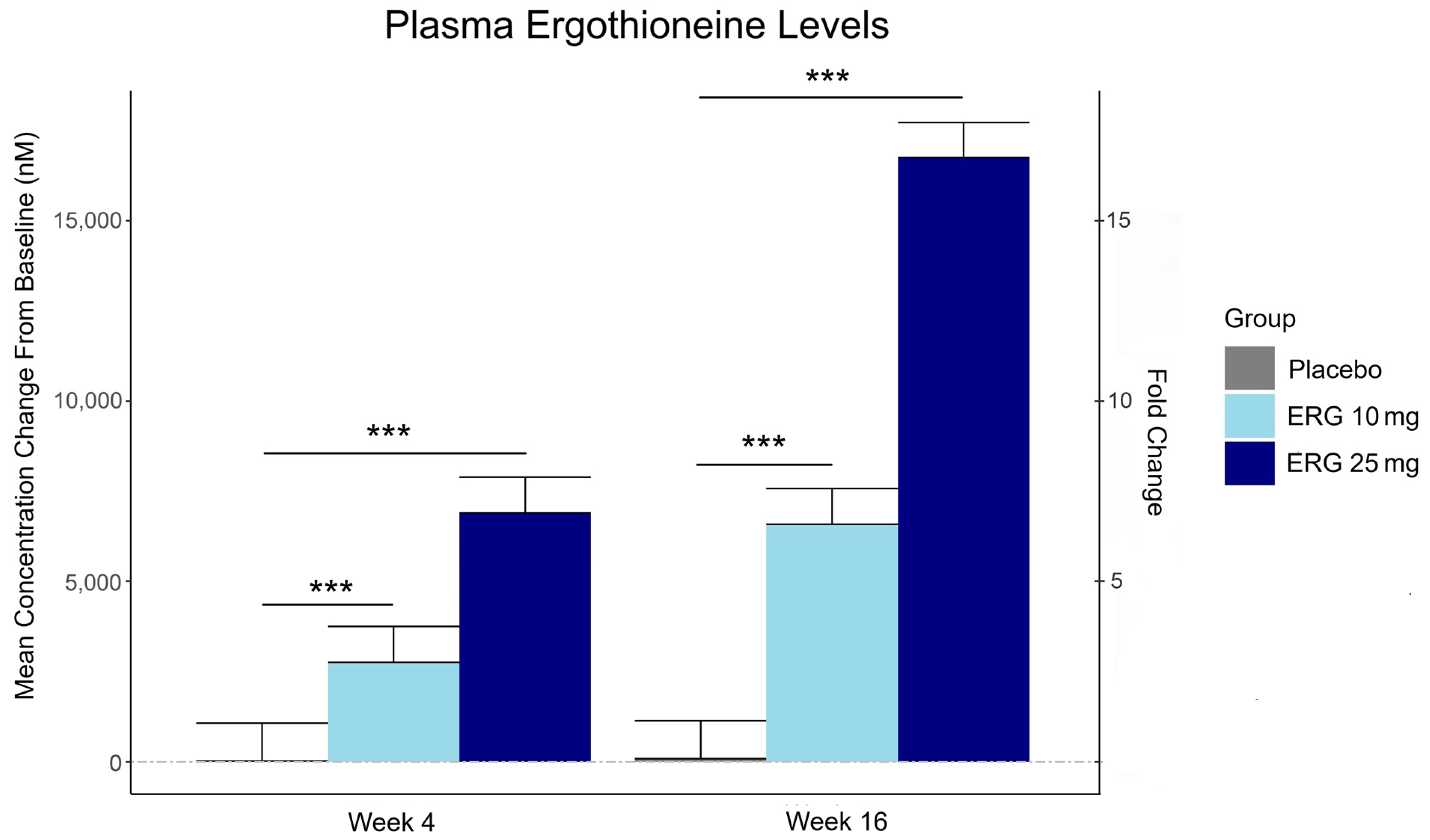
“In this 16-week randomized, double-blind, placebo-controlled trial, 147 adults aged 55–79 with subjective memory complaints received ergothioneine (10 mg or 25 mg/day ErgoActive®) or placebo. Across all the groups, approximately 73% of participants in each group were female, with a median age of 69 years.
The primary outcome was the change in composite memory. Secondary outcomes included other cognitive domains, subjective memory and sleep quality, and blood biomarkers. At baseline, participants showed slightly above-average cognitive function (neurocognitive index median = 105), with plasma ergothioneine levels of median = 1154 nM.
Although not synthesized in the human body, ergothioneine is efficiently absorbed via the OCTN1 transporter (also known as the ergothioneine transporter, or ETT), which is expressed in many tissues, including the intestine, red blood cells, kidneys, bone marrow, immune cells, skin, and brain. This transporter enables ergothioneine to accumulate in high concentrations in organs vulnerable to oxidative stress and inflammation. Ergothioneine has multiple cellular protective functions, including scavenging reactive oxygen species, chelating redox-active metals, suppressing pro-inflammatory signaling, and protecting mitochondrial function.
Plasma ergothioneine increased by ~3- and ~6-fold for 10 mg, and ~6- and ~16-fold for 25 mg, at weeks 4 and 16, respectively.
While the primary outcome, composite memory, showed early improvement in the 25 mg group compared to baseline, this effect was not sustained and did not differ from placebo. Reaction time showed a significant treatment-by-time interaction favoring ergothioneine, yet the between-group differences were not significant, suggesting that any potential benefits were modest and require validation in larger or longer studies.
Other cognitive effects observed were primarily within-group and not consistently dose-responsive, highlighting the challenge of detecting objective cognitive changes over a relatively short study duration in high-functioning healthy populations. However, positive effects of ergothioneine supplementation were observed on subjective measures of prospective memory and sleep initiation that were not seen in the placebo group.
This trial adds to the growing body of evidence supporting the favorable safety profile of ergothioneine. No adverse events attributable to ergothioneine were reported. Additionally, we observed potential hepatoprotective effects, with significant reductions in the plasma AST and ALT levels, particularly among males in the ERG 25 mg group.”
https://www.mdpi.com/1661-3821/5/3/15 “The Effect of Ergothioneine Supplementation on Cognitive Function, Memory, and Sleep in Older Adults with Subjective Memory Complaints: A Randomized Placebo-Controlled Trial”
The third graphic for Ergothioneine dosing, Part 2 showed a human study where a 25 mg dosing stopped after Day 7, but the plasma ergothioneine level stayed significantly higher than baseline through Day 35.
The second graphic for Ergothioneine dosing, Part 2 was a male mouse experiment where plasma ergothioneine levels of a human equivalent 22 mg to 28 mg daily dose kept rising through 92 weeks.
This trial couldn’t explain the desirability of a 25 mg daily dose that was likely (per the second and third graphics for Ergothioneine dosing, Part 2) to sustain the subjects’ increased plasma ergothioneine levels well after the trial ended at Week 16. What effects can be expected from a sustained plasma ergothioneine level that’s 16 times higher than the subjects’ initial levels? Were these 16-fold sustained plasma ergothioneine levels better or worse than the 6-fold increases in the 10 mg group, both of which were likely to continue past the trial’s end?
A representative of the trial’s sponsoring company talked a little more about the trial in this interview:
Another clinical trial investigated ergothioneine’s effects on skin:
“We conducted an 8-week, randomized, double-blind, placebo-controlled clinical trial to evaluate effects of daily oral supplementation with 30 mg of ergothioneine (DR.ERGO®) on skin parameters in healthy adult women aged 35–59 years who reported subjective signs of skin aging. Objective measurements including melanin and erythema indices, skin glossiness, elasticity, and wrinkle and pigmentation counts were used to comprehensively evaluate changes in skin condition.
The OCTN1 transporter is preferentially expressed in basal and granular epidermal layers, where cellular renewal and barrier maintenance are most active. Once internalized, ergothioneine localizes to mitochondria, where it directly scavenges reactive oxygen species (ROS) and protects mitochondrial DNA from UV- and inflammation-induced damage.
At the signaling level, ergothioneine activates key protective pathways such as the Nrf2/ARE axis, enhancing expression of antioxidant enzymes including HO-1, NQO1, and γ-GCLC. These enzymes contribute to redox homeostasis and glutathione regeneration, reinforcing cellular defense systems against photoaging and environmental insult.
In parallel, ergothioneine modulates the PI3K/Akt/Nrf2 and SIRT1/Nrf2 pathways, which are implicated in collagen preservation, inflammation resolution, and mitochondrial maintenance. These pathways converge to reduce matrix metalloproteinase (MMP) activity, enhance collagen synthesis, and suppress pro-inflammatory cytokines (TNF-α, IL-6, IL-1β), all of which are central to maintaining skin structure and function.
Compared to placebo, the DR.ERGO® ergothioneine group showed significantly greater improvements in melanin and erythema reduction, skin glossiness, elasticity, and wrinkle and spot reduction. No adverse events were reported.
These findings corroborate and extend previous clinical evidence from (Hanayama et al., 2024), who investigated an ergothioneine-rich mushroom extract (Pleurotus sp., 25 mg ergothioneine/day) in a 12-week randomized double-blind trial, and (Chunyue Zhang, 2023), who examined pure ergothioneine supplementation (25 mg/day) in a 4-week open-label study. We contextualized our results within this existing literature by comparing key outcomes.
Several limitations should be acknowledged:
The study cohort consisted solely of Japanese women aged 35–59 years, which may limit generalizability across sexes, ethnicities, and age groups.
The 8-week intervention period, while sufficient to detect short-term effects, does not allow conclusions about the sustainability of benefits or the risk of relapse upon discontinuation.
The placebo group also showed modest improvements in self-perception, highlighting the well-documented placebo response in beauty and wellness studies.
This study focused on a single daily dosage (30 mg/day) without evaluating dose–response relationships, and hydration-specific endpoints such as corneometry or transepidermal water loss (TEWL) were not included.”
Two clinical trials investigated ergothioneine’s effects on sleep quality:
“A four-week administration of 20 mg/day ergothioneine (EGT), a strong antioxidant, improves sleep quality; however, its effect at lower doses remains unclear. This study estimated the lower effective doses of EGT using a physiologically based pharmacokinetic (PBPK) model in two clinical trials.
In Study 1, participants received 5 or 10 mg/day of EGT for 8 weeks, and their plasma and blood EGT concentrations were measured. An optimized PBPK model incorporating absorption, distribution, and excretion was assembled. Our results showed that 8 mg/day of EGT for 16 weeks was optimal for attaining an effective plasma EGT concentration.
In Study 2, a randomized, double-blind, placebo-controlled study, participants received 8 mg/day EGT or a placebo for 16 weeks. The subjective sleep quality was significantly improved in the EGT group than in the placebo group.
In mammals, EGT is not generated in the body but is acquired from the diet via the carnitine/organic cation transporter OCTN1/SLC22A4. Its plasma concentration after oral administration is quite stable and gradually increases after repeated dosing on a multi-day basis.
Blood concentrations of EGT increase after Day 8 when EGT intake is interrupted, and they continue to increase until Day 35. The delayed increase in EGT concentration in the blood, compared with that in the plasma, can be interpreted as its efficient uptake by undifferentiated blood cells, which express high levels of OCTN1/SLC22A4 in the bone marrow, and subsequent differentiation to mature blood cells that enter the circulation. This may imply the nonlinear absorption, distribution, and excretion of EGT owing to saturation of the transporter at higher concentrations, potentially leading to difficulty in model construction.
This is the first study to propose a strategy to estimate lower effective doses based on the PBPK model.”
The bolded section above referenced a 2016 study / third graphic for Ergothioneine dosing, Part 2, where a 25 mg dosing stopped after Day 7, but the plasma ergothioneine level stayed high through Day 35. I didn’t see that the referenced 2004 and 2010 studies addressed this 2016 finding.
I also didn’t see that this study’s mathematical model accounted for saturation of the OCTN1 transporter or other effects, such as a very small ergothioneine clearance rate. Okay, lower the ergothioneine dose, and achieve a lower persistent plasma ergothioneine level, to what benefit?
“The present study demonstrated that OCTN1 is associated with myeloid cells rather than lymphoid cells, and especially with erythroid-lineage cells at the transition stage from immature erythroid cells to peripheral mature erythrocytes.”
Persistent high ergothioneine levels aren’t costless. Skewing bone marrow stem cells and progenitor cells toward a myeloid lineage is done at the expense of lymphocytes, T cells, B cells, and other lymphoid lineages.
Where are the studies that examine these tradeoffs? Subjective sleep quality in this study and sleep initiation in the first study above aren’t sufficiently explanatory.
A study investigated associations of plasma ergothioneine levels and cognitive changes in older adults over a two-year period:
“Observational studies have found that lower plasma levels of ergothioneine (ET) are significantly associated with higher risks of neurodegenerative diseases. However, several knowledge gaps remain:
Most of the above studies were based on cross-sectional study design, and potential reverse causation cannot be excluded. It has been suggested that plasma ET declines concomitantly with the deterioration of cognitive function.
Since the impact of a single dietary factor on health is mild, it is prone to be affected by the baseline characteristics of subjects (such as sex, educational level, disease status and gene polymorphism). However, no study has systematically evaluated potential effect modifiers on the association between ET levels and cognitive function.
The dose-response distribution between ET and cognitive function remains undetermined.
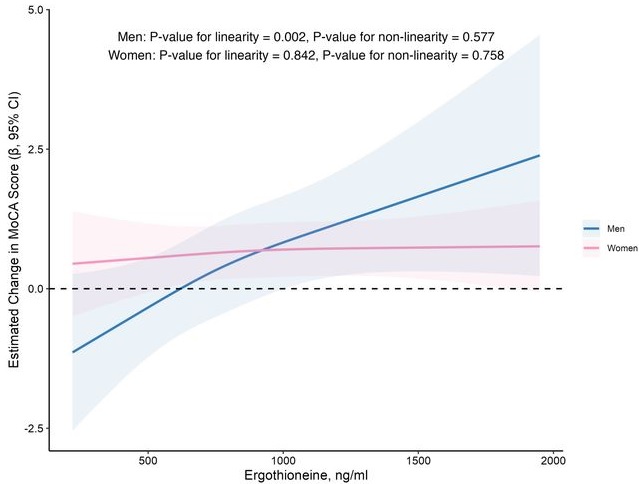
In this prospective cohort study of 1,131 community-dwelling older adults (mean age 69 years), higher baseline plasma ET levels were significantly associated with slower cognitive decline, as assessed by Montreal Cognitive Assessment (MoCA) scores, during a 2-year follow-up period.
When the plasma concentration of ET exceeds 1,000 ng/mL, the decline in cognitive function significantly slows down. However, this association has only been observed in men.
Domain-specific analysis found that the observed ET-MoCA association was mainly driven by the temporary slowdown in the decline of visuospatial/executive and delayed recall. Impaired delayed recall represents one of the earliest and most sensitive cognitive markers of dementia progression, predictive of conversion from MCI to dementia. The preferential preservation of this function by ET suggests targeted neuroprotective effects within the hippocampus.
Visual inspection of the spline curves revealed a potential plateauing effect at ET concentrations ≥1,000 ng/mL in the total population.
Baseline ET concentrations differed between men and women. Most men (81.5%) had concentrations below 1,000 ng/mL (median 754.2, IQR 592.0–937.9 ng/mL). Women exhibited substantially higher median plasma ET concentrations than men, with 35.7% of women exceeded 1,000 ng/mL (median 890.1, IQR 709.7–1,095.6 ng/mL).
Our study included only participants with normal cognitive function, and the results remained robust even after excluding those with baseline cognitive function at the lower end of the normal range. Collectively, our findings support that low ET intake occurs prior to cognitive decline.
Our findings indicate that higher plasma ET levels are significantly associated with slower cognitive decline independent of confounders in non-demented community-dwelling elderly participants, with such association observed in men but not women. Dose-response curves indicated plateauing effects above 1000 ng/mL.”
The average age of this study and the first trial above were both 69 years. Since the first trial’s participants showed slightly above-average cognitive function (neurocognitive index median = 105), with plasma ergothioneine levels of median = 1154 nM at baseline, and this study showed plateauing effects above 1000 ng/mL, I wonder how raising plasma ergothioneine levels above 1000 ng/mL could possibly show a net benefit for older people? What are the trade-offs for older people between potentially increasing slightly above-average cognitive function with ergothioneine and its other effects from saturating their OCTN1 transporter?
This study is at its preprint stage. I’m interested to see if its peer review prompts these researchers to also investigate the common finding that people who are most deficient at baseline have the greatest improvements. If so, would these sex-specific associations still hold?
Wrapping up with a study that investigated associations of serum ergothioneine levels with the risk of developing dementia:
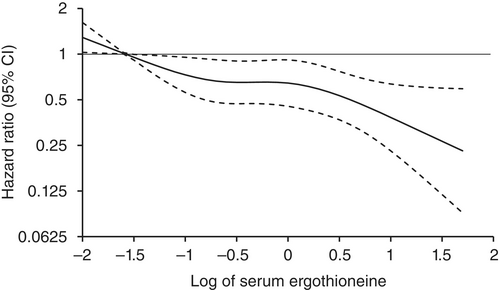
“1344 Japanese community-residents aged 65 years and over, comprising 765 women and 579 men, without dementia at baseline were followed prospectively for a median of 11.2 years.
During follow-up, 273 participants developed all-cause dementia. Among them, 201 had Alzheimer’s disease (AD) and 72 had non-Alzheimer’s disease (non-AD) dementia.
Age- and sex-adjusted hazard ratios (HRs) for all-cause dementia, AD, and non-AD dementia decreased progressively across increasing quartiles of serum ergothioneine. These associations remained significant after adjustment for a wide range of cardiovascular, lifestyle, and dietary factors, including daily vegetable intake.
In subgroup analysis, association between serum ergothioneine levels and the risk of dementia tended to be weaker in older participants and in women:
In older individuals, cumulative burden of multiple risk factors such as hypertension, diabetes mellitus, and smoking may contribute to both neurodegenerative and vascular pathology, potentially diminishing the relative influence of ergothioneine.
In women, postmenopausal hormonal changes, particularly the decline in estrogen, have been associated with increased oxidative stress and a higher vulnerability to neurodegenerative changes.
Several limitations should be noted:
Since serum ergothioneine levels and other risk factors were measured only at baseline, we could not evaluate the changes of serum ergothioneine levels during the follow-up period. Lifestyle modifications during follow-up could have influenced serum ergothioneine levels and other risk factors. In addition, serum ergothioneine level was measured only once, and from a sample.
We cannot rule out residual confounding factors, such as other nutrients in mushrooms and socioeconomic status.
There is a possibility that dementia cases at the prodromal stage were included among participants with low serum ergothioneine levels at baseline.
We are unable to specify which mushroom varieties were predominantly consumed by participants in the town of Hisayama.
Given the limited discriminative ability of serum ergothioneine and potential degradation due to long-term sample storage, we were unable to explore a clinically meaningful threshold value of serum ergothioneine.
Generalizability of findings was limited because participants of this study were recruited from one town in Japan.
These findings suggest that the potential benefit of ergothioneine may be attenuated in individuals with pre-existing, multifactorial risk profiles for dementia.
Our findings showed that higher serum ergothioneine levels were associated with a lower risk of developing all-cause dementia, AD, and non-AD dementia in an older Japanese population. Since ergothioneine cannot be synthesized in the human body, a diet rich in ergothioneine may be beneficial in reducing the risk of dementia.”
For five years I got most of my estimated 7 mg daily ergothioneine intake from mushrooms in AGE-less chicken vegetable soup per Ergothioneine dosing. The soup was always boring, but I got too bored this year and stopped making it. I haven’t replaced mushroom intake with supplements.
I still don’t eat fried or baked foods, preferring sous vide and braising cooking methods to avoid exogenous advanced glycation end products. I avoid buying foods that evoke a hyperglycemic response or otherwise form excessive endogenous AGEs per All about AGEs.
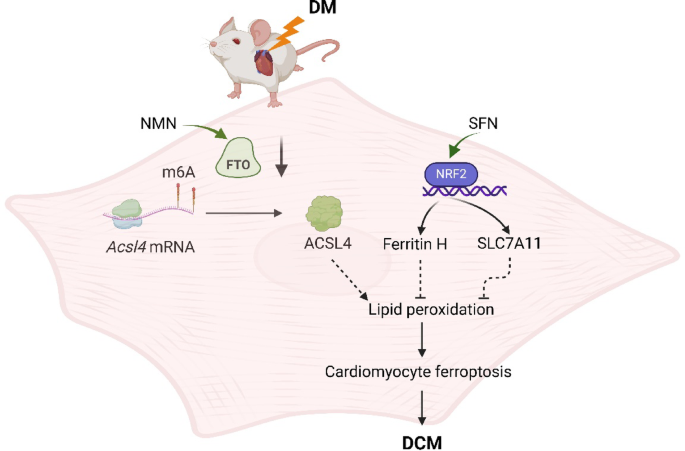
A 2025 rodent study investigated synergistic effects of sulforaphane (SFN) and nicotinamide mononucleotide (NMN) on diabetic cardiomyopathy:
“Diabetic cardiomyopathy (DCM) as a significant diabetes complication remains a major human challenge. In this study, we provide evidence that the fat mass and obesity-associated protein (FTO) plays a pivotal role in DCM pathogenesis.
Downregulation of FTO in DCM acts as a critical inducer of ferroptosis by increasing expression of acyl-CoA synthetase long-chain family 4 (ACSL4), a key positive mediator of ferroptosis. FTO-mediated mitigation of ferroptosis occurs in an ACSL4-dependent manner which leads to increased methylation of Acsl4 transcripts.
Ferroptosis plays an essential role in the pathogenesis of DCM.
As the most widespread mRNA modification, N6-methyladenosine (m6A) is globally downregulated and implicated in diabetes and its complications.
FTO, which is an m6A demethylase, was found to be downregulated in diabetes and its cardiovascular complications.
NAD+ enhances the demethylase activity of FTO. Dietary supplementation with NMN, a critical intermediate in the NAD+biosynthetic pathway, has been shown to efficiently elevate endogenous NAD+ levels.
Enhancing the demethylase activity of FTO with NMN combined with SFN targeting NRF2 could synergistically reduce the level of lipid peroxides to inhibit ferroptosis, providing an effective avenue for alleviating DCM.
We found that NMN could alleviate ferroptosis and improve heart function through enhancing FTO. SFN could prevent ferroptosis and partly rescue heart function via AMPK-mediated NRF2 activation.
We demonstrated that SFN combined with NMN treatment could significantly inhibit lipid peroxidation and rescue cardiac function in DCM compared to SFN or NMN treatment alone.
Although the combined regimen further suppressed ferroptosis and improved cardiac performance, it fell short of complete remission, underscoring that additional pathways also contribute substantially to the pathogenesis of DCM.”
The epigenetic mechanism involved with this study’s dietary dissolved-in-water 100mM NMN dose was Non-CpG methylation. This study used the same very low sulforaphane dose intraperitoneally injected as Eat broccoli sprouts for your heart. Discussion of that study provided an example that if a person waited until a diabetes-related disease condition became a problem, capabilities to adequately address causes and prevent the problem may be lost.
Notice in the last bar of the second graphic above taken from Figure 7 that the combined treatment was also provided to non-diabetic mice. These researchers provided over a dozen other measurements in Figure 7 to show similar short-term non-effects of the combined treatment, i.e. that it neither benefited nor harmed non-diabetic subjects. Grok interpreted this study’s 3-month-long intervention to be a 1-to-5 year human equivalent, depending on the measured effect (shorter for metabolic effects like MDA, longer for structural cardiac changes like reduced ferroptosis.)
The male subjects began at 2-months old, a human-equivalent 15-20 years old. These researchers gave them diabetes by feeding them a “high-fat diet for 3 months to induce insulin resistance, followed by a single intraperitoneal injection of streptozotocin (STZ) (in 0.1 mol/L of citrate acid buffer, 60 mg/kg) to induce partial insulin deficiency.” A 5-months old mouse is a 25-30 years old human equivalent.
Grok considered this study’s NMN human equivalent dose to be extremely high if provided in drinking water, but not if injected, depending on volume. However, the study didn’t state that its NMN dose was injected, and there was no dose volume indicated.
Here are two 2025 papers, starting with a rodent study that investigated interactions between the Nrf2 and kynurenine pathways:
“Exposure to the tryptophan metabolite kynurenine and its electrophilic derivative kynurenine-carboxyketoalkene (Kyn-CKA) leads to an increase in the abundance of transcription factor Nrf2 and induction of Nrf2-target genes. The Keap1/Nrf2 system is the main orchestrator of cellular defence against environmental stress, most notably oxidative and inflammatory stress.
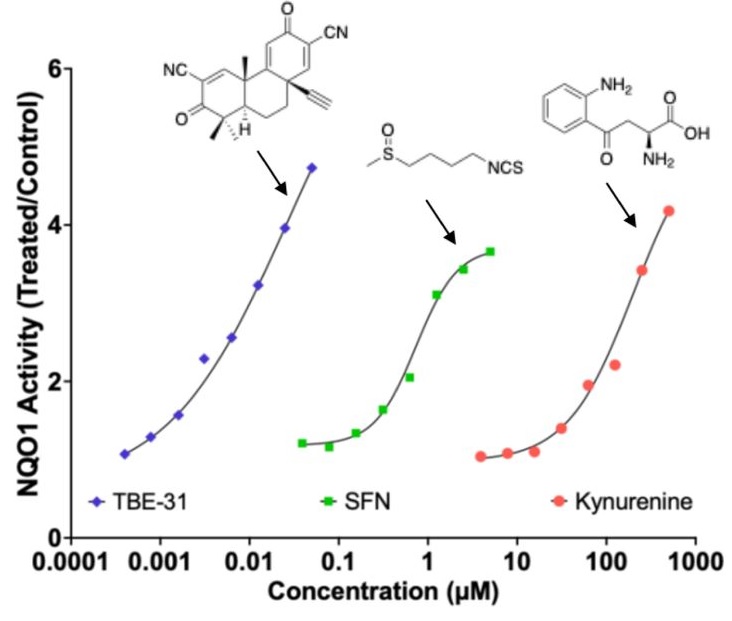
Nrf2 can be activated pharmacologically by small molecules, the majority of which are electrophiles and oxidants that modify specific cysteine-based sensors in Keap1. C151 in Keap1 is the target of the isothiocyanate sulforaphane, a classical Nrf2 activator that has been employed in ∼90 clinical trials, as well as for the two Nrf2 activators that are clinically in use: dimethyl fumarate, for relapsing remitting multiple sclerosis, and omaveloxolone, for Friedreich’s ataxia.
Kynurenine is an endogenous metabolite derived from the essential amino acid tryptophan. Kynurenine and its metabolites, such as the electrophilic kynurenine-carboxyketoalkene (Kyn-CKA), have been demonstrated to activate Nrf2 in other pathologies, including sickle cell disease, attenuating inflammation. Moreover, identification of the gene encoding the kynurenine-metabolising enzyme kynureninase as a gene transcriptionally upregulated by Nrf2, provides a plausible negative feedback regulatory mechanism.
Because kynurenine is not electrophilic, whereas its metabolite Kyn-CKA is, we considered the possibility that Kyn-CKA is the actual Nrf2 activator. Using biochemical and cell-based assays, we found that Kyn-CKA reacts with C151 in the BTB domain of Keap1 and increases the thermostability of Keap1, indicating target engagement. Consequently, Nrf2 accumulates and induces transcription of antioxidant/electrophile-responsive element (ARE/EpRE)-driven genes.
These findings demonstrate that Kyn-CKA targets C151 in Keap1 to derepress Nrf2, and reveal that Nrf2 is a main contributor to the anti-inflammatory activity of Kyn-CKA in macrophages.”
A review subject was targeting nicotinamide adenine dinucleotide, oxidized form (NAD+) for clinical use:
“Mammalian NAD+ biosynthesis includes four known pathways, primarily occurring in cytoplasm:
(a) the NRH pathway;
(b) the salvage pathway;
(c) the Preiss–Handler pathway; and
(d) the kynurenine pathway.
The de novo kynurenine pathway metabolizes tryptophan (Trp) to NAD+, producing various intermediates that serve as biomarkers for different diseases. These intermediates show alterations in various pathological conditions.
While kynurenine and its metabolic derivatives are intermediates in the de novo NAD+ biosynthesis pathway, these are also produced independently in various physiological contexts, particularly in immune cells, where they act as immunomodulatory compounds.”
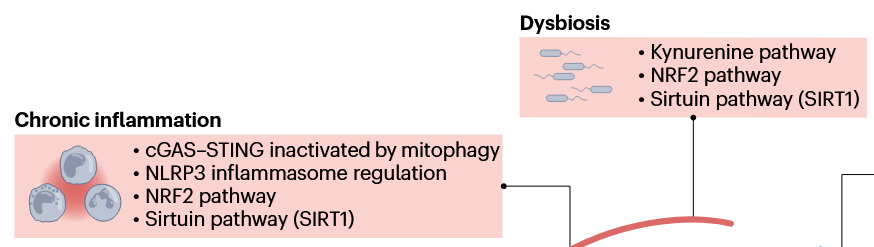
This second paper above showed a graphic of the Nrf2 and kynurenine pathways together in a diagram showing relationships between NAD+ augmentation and the hallmarks of aging, but didn’t elaborate other than labeling their box Dysbiosis. So how these two pathways interact is better outlined in the first paper above with explaining how a kynurenine-metabolizing enzyme is one of the hundreds of Nrf2 target genes, creating a natural feedback loop between Nrf2 activation and the kynurenine pathway.
These reviewers also lumped SIRT1 in their Dysbiosis box, and into several other boxes, probably due to the penultimate coauthor’s influence:
However, repeating something over and over doesn’t make it scientifically valid regardless of the number of citations. Or, as a 2022 review Sirtuins are not conserved longevity genes concluded:
“A global pursuit of longevity phenotypes was driven by a mixture of framing bias, confirmation bias, and hype. Review articles that propagate these biases are so rampant that few investigators have considered how weak the case ever was for sirtuins as longevity genes.
Acknowledging that a few positive associations between sirtuins and longevity have been identified after thousands of person-years and billions of dollars of effort, we review the data and suggest rejection of the notions that sirtuins (i) have any specific connection to lifespan in animals and (ii) are primary mediators of the beneficial effects of NAD repletion.”
Continuing Part 1, here are four more 2025 human studies of the Nrf2 activator astaxanthin, starting with a randomized, double-blind, placebo-controlled trial of its effects on reducing oxidative stress and inflammatory responses following eccentric exercise:
“This study investigated effects of astaxanthin supplementation on plasma MDA and HMGB1 levels following acute eccentric exercise in recreationally active male students. Fifty-four students were assigned to receive either 12 mg/day of natural astaxanthin (AST, n = 27) or placebo (PLA, n = 27) for 14 days.
A key consequence of eccentric-induced muscle damage is overproduction of reactive oxygen species (ROS). When ROS production exceeds the capacity of endogenous antioxidant systems, lipid peroxidation can occur. Malondialdehyde (MDA) is a stable end-product of lipid peroxidation and serves as a widely recognized biomarker for oxidative stress and cell membrane damage.
In parallel, muscle cell damage results in release of damage-associated molecular patterns (DAMPs) into the extracellular space. Among these, High Mobility Group Box-1 (HMGB1) plays a central role in inflammation when passively released from the nucleus. HMGB1 acts as a potent pro-inflammatory signal by activating innate immune receptors, recruiting immune cells, and upregulating cytokines such as IL-6 and TNF-α.
This heightened immune activity contributes to delayed-onset muscle soreness, which typically peaks 24–72 hours post-exercise, and is associated with impaired recovery. Sustained elevations in oxidative and inflammatory biomarkers, including MDA and HMGB1, may further impair recovery and contribute to long-term muscle pathology.
Astaxanthin’s antioxidant effects are mediated through both direct and indirect mechanisms. Structurally, astaxanthin is a xanthophyll carotenoid with a unique polar–nonpolar–polar configuration that enables it to span the phospholipid bilayer of cell membranes. This positioning allows it to neutralize ROS both at the membrane surface and within the lipid bilayer.
In addition, astaxanthin enhances endogenous antioxidant defenses by upregulating enzymes such as superoxide dismutase (SOD), catalase, and glutathione peroxidase (GPx) through activation of the Nrf2–ARE signaling pathway. This dual mode of action provides both immediate and sustained protection against oxidative stress during and after exercise.
The placebo group showed substantial increases in MDA and HMGB1 after exercise, whereas the astaxanthin group experienced attenuated rises (~22% and ~27% smaller, respectively) and faster recovery toward baseline within 24 hours. These findings suggest that astaxanthin supplementation can be incorporated into recovery strategies for athletes and active individuals, especially during periods of heavy training or repeated bouts of intense eccentric exercise. By reducing oxidative damage and inflammation, astaxanthin may shorten recovery time, limit performance loss, and support overall training adaptations—benefits that are particularly valuable in sports requiring frequent high-intensity efforts.
Several limitations should be acknowledged in this study.
Sample size was relatively small and limited to recreationally active young males, which may restrict generalizability of findings to other populations such as females, older adults, or elite athletes.
Supplementation period was limited to 14 days; although this duration is sufficient to achieve plasma saturation of astaxanthin, longer interventions may produce different or more pronounced effects.
Only two biomarkers were assessed (MDA and HMGB1), which provide important but incomplete insights into broader oxidative stress and inflammatory response. Including additional markers such as enzymatic antioxidants, cytokine profiles, and muscle damage indicators (e.g., creatine kinase) could yield a more comprehensive understanding.
Dietary intake and physical activity outside the intervention were self-reported and not strictly controlled, which may have introduced variability in results.”
https://tmfv.com.ua/journal/article/view/3664/1922 “Taking Astaxanthin Supplementation Attenuates MDA and HMGB1 Following Eccentric Exercise: A Randomized Controlled Trial in Recreationally Active Students”
A clinical trial investigated astaxanthin’s effects with exercise in diabetic women:
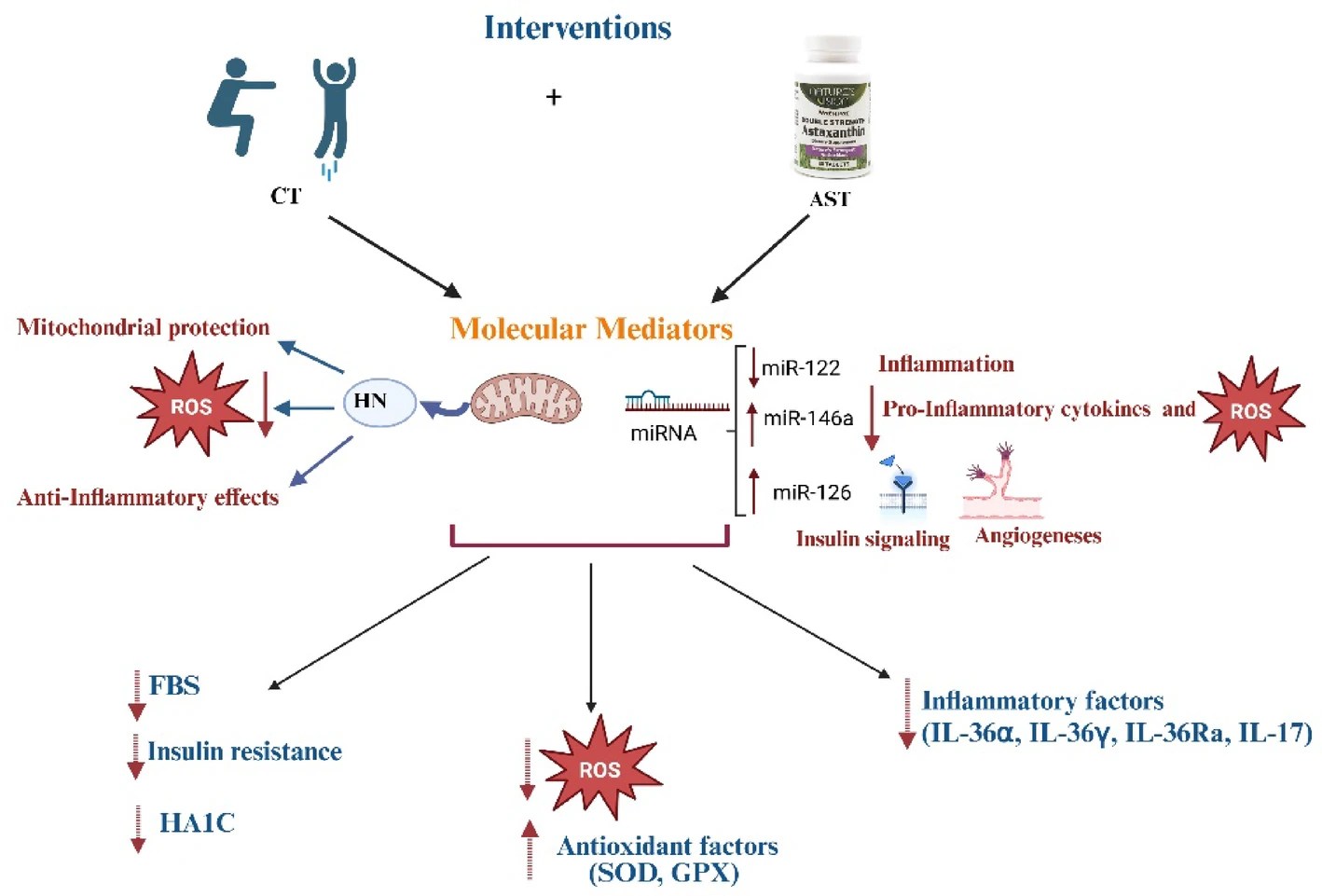
“This study examined whether combined aerobic and resistance training (CT) and astaxanthin (AST) supplementation synergistically improve oxidant and inflammatory status as well as metabolic indices in T2DM, focusing on the mediatory role of Humanin (HN) and microRNAs (miRNA-122, miRNA-126-3p, and miRNA-146a).
Ninety women with T2DM were randomly assigned to six groups (n = 15 each):
Control (C), placebo (P), AST supplementation (S), combined training (CT), CT + placebo (CT + P), and CT + AST supplementation (CT + S).
CT, CT + P and CT + S groups underwent an 8-week training program (eight exercises, three sessions per week).
S and CT + S groups received 8 mg/day of AST.
This study only enrolled female participants age between 30 and 60 years old to minimize inter-individual biological variability arising from sex differences in hormone regulation, fat distribution, and gene expression related to inflammation and oxidative stress. Oxidative stress (OS) markers, inflammatory cytokines, HN levels, miRNAs expression, fasting blood glucose (FBG), insulin resistance (HOMA-IR), lipid profile, and hemoglobin A1c (HbA1c) were assessed.
HN is a member of a class of novel mitochondrial-derived peptides released during mitochondrial dysfunction. HN reduces ROS production, enhances antioxidant protein expression, maintains redox balance, and suppresses TNF-α, IL-1β, and IL-6 to inhibit inflammation. Furthermore, resistance and endurance training has shown to increase HN expression in patients with prediabetes. Exercise – aerobic and endurance – has been shown to increase circulating and skeletal muscle levels of HN, correlating with improved insulin sensitivity and mitochondrial function.
Our results showed:
CT and AST supplementation both improved antioxidant defense and reduced inflammation, and their combination was more effective than either intervention alone.
CT and AST supplementation increased blood concentration of HN, and their combination showed greater effects than AST supplementation, but not CT.
CT and AST supplementation increased blood levels of miRNAs-126-3p, and -146a and decreased miRNA-122, with their combination being slightly more effective in decreasing miRNA-122.
Both interventions improved lipid profile, with their combination being more effective in improving HDL and TG levels, although not total cholesterol.
FBG, HOMA-IR, and HbA1c were reduced by CT but not by AST supplementation.
Our data suggest that combining exercise with AST supplementation might improve oxidative status and inflammation through mechanisms involving HN and miRNAs 122, 126-3p, and 146a. Alleviating OS and inflammation could, in turn, lead to improvements in lipid profiles (e.g., TG, and HDL), IR, and reductions in HbA1c and FBG, as observed in our study. Furthermore, the combined approach seems to be more effective at improving cholesterol and TG levels.“
A meta-analysis of randomized controlled trials reported until May 2025 assessed astaxanthin’s effects on lipid profiles. Neither of the two trials covered here nor the three trials covered in Part 1 were included in this meta-analysis.
“Astaxanthin, a xanthophyll carotenoid, has garnered significant interest due to its benefits with regard to dyslipidemia. This multifaceted functional food ingredient modulates several key enzymes associated with lipid regulation, including HMG-CoA reductase, CPT1, ACCβ, and acyl-CoA oxidase. It influences key antioxidant molecular pathways like Nrf2, limiting dyslipidemia occurrence and regulating liver cholesterol uptake through modulation of liver lipid receptors.
Astaxanthin daily doses and durations of analyzed studies: 12 mg for 8 weeks; 12 mg for 4 weeks; 20 mg for 12 weeks (two trials); 12 mg for 12 weeks; 8 mg for 8 weeks; 6 mg and 12 mg for 12 weeks; 6 mg, 12 mg, and 18 mg for 12 weeks.
This meta-analysis concludes positive effects of astaxanthin (6–20 mg/d) on HDL-C and triglyceride levels. Astaxanthin (6–20 mg/d) does not appear to significantly influence LDL-C and total cholesterol levels.
Regarding HDL-C, improvements were observed from 55 ± 8 mg/dL (pre-intervention) to 63 ± 8 mg/dL (post-intervention) (p < 0.01) in the 12 mg/d of astaxanthin groups. In triglyceride levels, results show a decrease from 151 ± 26 mg/dL (pre-intervention) to 112 ± 40 mg/dL (post-intervention) (p < 0.01) for 18 mg/d astaxanthin supplementation.
Further research is necessary to fully harness the potential of astaxanthin, which includes assessing astaxanthin in different subsets of patients, and in combination with other nutraceuticals to understand the compound’s effectiveness with regard to varying health conditions, genetic and epigenetic factors, and synergistic effects with other compounds.”
https://www.mdpi.com/1424-8247/18/8/1097 “Assessing the Effects of Moderate to High Dosage of Astaxanthin Supplementation on Lipid Profile Parameters—A Systematic Review and Meta-Analysis of Randomized Controlled Studies”
This same group of researchers assessed that in nine RCTs, astaxanthin had no effects on either body weight or BMI per https://www.mdpi.com/1424-8247/18/10/1482 “Therapeutic Potential of Astaxanthin for Body Weight Regulation: A Systematic Review and Meta-Analysis with Dose–Response Assessment”
Here are three 2025 clinical trials of the Nrf2 activator astaxanthin’s effects. Let’s start with a clinical trial of inflammation-related diabetic complications and insulin resistance:
“We investigated effects of 10 mg/day astaxanthin (ASX) supplementation for 12 weeks on microRNAs (miRNAs), lysophosphatidylcholine (LPC), and α-hydroxybutyrate (α-HB) as novel factors in development of a variety of diabetes-related complications.
LPC is believed to play a significant role in atherosclerosis and inflammatory diseases by modifying functions of multiple cell types, including smooth muscle cells, endothelial cells, monocytes, macrophages, and T cells. LPC can interfere with glucose-stimulated insulin secretion by impairing calcium homeostasis and other signaling pathways that are crucial for the proper functioning of beta cells. This impairment exacerbates hyperglycemia in diabetic patients. LPCs may impede insulin signaling pathways, thereby contributing to insulin resistance (IR).
α-HB is also an indicator of IR and impaired glucose regulation, both of which appear to result from excessive lipid oxidation and oxidative stress. The European population cohorts in 2016 identified α-HB as a selective biomarker for decreased glucose tolerance and prediabetes, which was independent of age, sex, BMI, and fasting glucose.
A number of studies have established a link between miR-21, miR-34a, and miR-155 and diabetic complications such as retinopathy and nephropathy.
In the ASX group, participants were divided into 2 subgroups according to the urinary albumin-to-creatinine ratio (ACR) (< 30 mg/g or ≥ 30 mg/g, an indicator of diabetic kidney disease).
The level of fasting plasma glucose before and after 12 weeks of treatment with ASX was 139.27 ± 21.18 vs. 126.43 ± 18.97 (p = 0.002), demonstrating a significant reduction compared to the placebo group.
In the ASX group, the mean HbA1c level at baseline was 7.89 ± 0.79 and declined to 7.05 ± 0.35 after the supplementation period, which was statistically significant.
Supplementation with ASX resulted in a statistically significant drop in HOMA-IR levels, whereas this parameter was not altered significantly in the placebo group.
The ASX group, in comparison with the placebo group, demonstrated marked changes in lipid profile factors such as TC, TG, and LDL (p = 0.011, p = 0.043, and p = 0.022, respectively).
Clinical studies indicate that rigorous diabetes management does not substantially diminish appearance of complications. Modifications in oxidative stress and IR markers, as well as miRNA expression, must be analyzed to identify biological markers with sufficient predictive power for development of complications in diabetic patients.
Supplementation with ASX substantially diminished the levels of α-HB, LPC, and inflammation-related miRNAs in diabetic patients with and without complications.”
Another clinical trial investigated astaxanthin’s effects in heart failure patients:
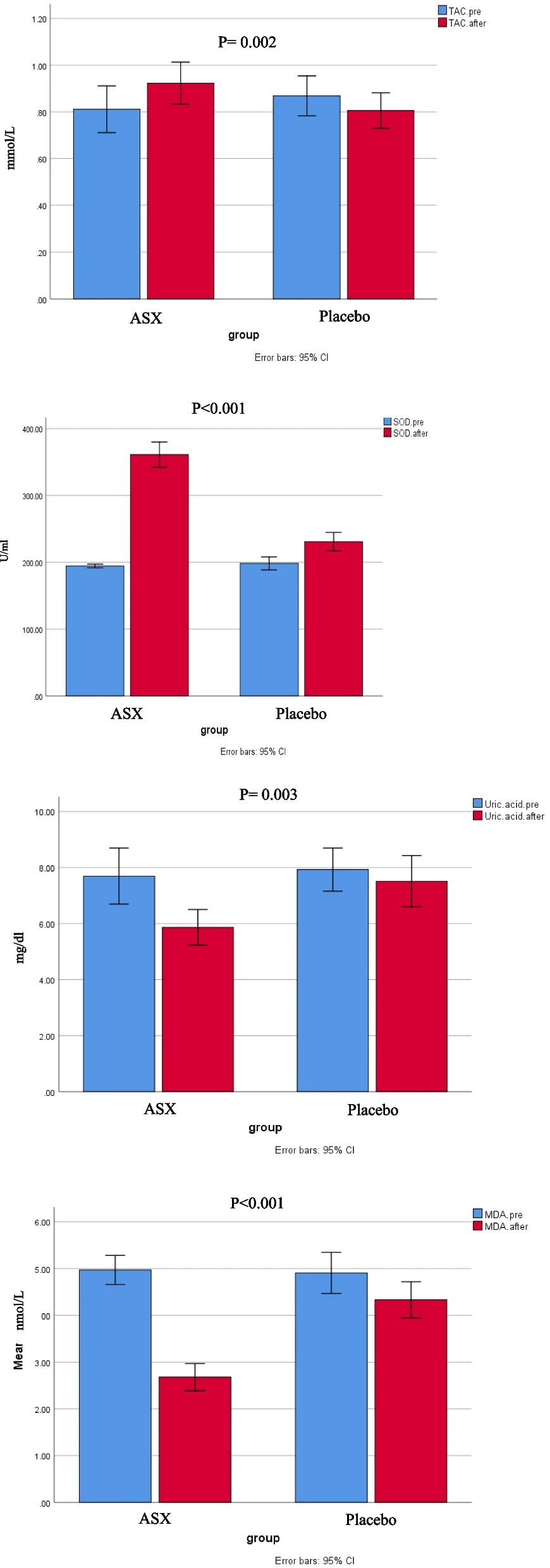
“Chronic heart failure (HF) is often linked to increased oxidative stress and metabolic issues like high uric acid, which can worsen outcomes.This study aimed to investigate the effects of ASX supplementation on oxidative stress markers as the primary outcome and clinical symptoms in patients with HF.
80 patients with HF were enrolled and randomly assigned to receive either ASX (20 mg/day) or a placebo (20 mg/day of maltodextrin) for 8 weeks. Biomarkers including total antioxidant capacity (TAC), malondialdehyde (MDA), superoxide dismutase (SOD), serum uric acid (UA), and clinical symptoms (dyspnea, fatigue, appetite) were assessed pre-and post-intervention.
Daily supplementation with 20 mg of ASX for eight weeks in patients with HF resulted in significantly greater improvements in oxidative stress biomarkers compared to placebo group. This improvement included reductions in uric acid and MDA, along increases in TAC and SOD.
In our study, participants received the cis-isomer form of ASX. The cis-isomer of ASX demonstrates greater anti-inflammatory and antioxidant properties than the trans-isomer, along with enhanced bioavailability. Inconsistencies among studies may be attributed to differences in participants’ baseline antioxidant status, underlying medical conditions, dosage, isomeric form and formulation of ASX used, and the duration of intervention.
One of the strengths of this study is that it represents the first randomized clinical trial to evaluate the effects of ASX supplementation on oxidative stress markers, UA levels, and clinical symptoms in patients with HF. Additionally, potential confounding factors were controlled as much as possible. However, several limitations were identified, including the relatively short intervention duration, limited sample size, limited generalizability of the findings due to the single-center design, absence of blood ASX level measurements, and lack of long-term follow-up.”
A third clinical trial evaluated astaxanthin’s effects as an adjunct to standard treatment of community-acquired pneumonia:
“Adult patients diagnosed with community-acquired pneumonia (CAP) were enrolled and assigned to receive either 12 mg/day ASX or a placebo in addition to standard antibiotic therapy for 7 days. Inflammatory markers, including interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), and interleukin-10 (IL-10), were measured at baseline and post-treatment. Secondary outcomes included Sequential Organ Failure Assessment (SOFA) and Acute Physiology and Chronic Health Evaluation II (APACHE II) scores.
A total of 80 patients (40 per group) completed the study. Patients receiving ASX exhibited significant reductions in pro-inflammatory cytokines compared to the placebo group. IL-6 and TNF-α levels were significantly lower in the ASX group at the end of the study (P < 0.05). Additionally, SOFA and APACHE II scores showed greater improvements in ASX-treated patients, suggesting a potential role in mitigating disease severity.
These findings suggest that ASX may help preserve organ function, limit the progression of inflammatory injury, and reduce overall disease severity in hospitalized patients with CAP.
ASX is widely regarded as the most potent carotenoid, owing to its unique molecular structure. Its polar-nonpolar-polar configuration enables it to span lipid bilayers and neutralize ROS both within and outside cellular membranes—an advantage not shared by other carotenoids that tend to localize at the membrane surface.
Despite the positive findings of this study, some limitations should nevertheless be considered.
The relatively small sample size may have limited the statistical power to detect differences in some outcomes and affects the generalizability of the findings.
Microbiological data on CAP pathogens were not collected. As different microorganisms can trigger distinct inflammatory responses, this limits our ability to assess pathogen-specific variations in ASX efficacy.
A notable limitation of this study is the short follow-up duration, with outcomes assessed only over a 7-day period. While this timeframe offers insight into the acute effects of ASX on inflammatory and OS markers, it does not clarify whether these benefits are sustained beyond the immediate treatment window.
The fixed dose of 12 mg once daily may not have maintained optimal therapeutic levels throughout the day. Dose-ranging studies and evaluations of alternative regimens are needed to determine the most effective strategy.”
Wrapping up Plasmalogens Week with a summary of my plasmalogen-related experiences over the past two years since Plasmalogens, Part 3 in November 2023.
I took detailed plasmalogen measurements on July 24, 2025, with Dr. Goodenowe’s BioScan product. I’d guess that the populations against which BioScan personal Z-scores are derived are from Dr. Goodenowe’s research during this century, many frozen samples of which he’s kept. If so, I’d guess that these populations’ data probably don’t have bell-shaped curves, and that their data’s means and standard deviations may be skewed as they’re representing people who were diseased and/or old.
Here’s my peroxisomal function panel:
I wasn’t taking ProdromeNeuro or ProdromeGlia at the BioScan blood draw time. ProdromeNeuro and ProdromeGlia supplements contain plasmalogen precursors that bypass peroxisome organelles’ normal plasmalogen synthesis functions. I haven’t reordered these supplements in 2025, but took them until my supplies ran out in January 2025. Don’t know to what extent their effects may have continued for six months.
Every day for months before the BioScan, I took a fish oil capsule with 690 mg EPA and 310 mg DHA, and a flax seed oil capsule (700 mg alpha linolenic acid omega-3, 154 mg linoleic acid omega-6, and 168 mg oleic acid omega-9). I also ate 3 eggs a day.
These practices influenced the above peroxisomal function results. My Z-scores of DHA and EPA ethanolamine plasmalogens (DHA +1.3, EPA +1.7) are more than one standard deviation above their respective population means.
The next step of plasmalogen synthesis after peroxisomes takes place in endoplasmic reticulum organelles. Among other papers describing these steps in the ER link’s results, Improving peroxisomal function states:
“Proper functioning of peroxisomes in metabolism requires the concerted interaction with other subcellular organelles, including the endoplasmic reticulum (ER), mitochondria, lipid droplets, lysosomes, and the cytosol. A striking example of peroxisome-ER metabolic cooperation is de novo biosynthesis of ether phospholipids.”
ER stress involves the unfolded protein response, a protein homeostasis-maintaining system that monitors ER conditions by sensing inadequacy in ER protein folding capacity. ER stress is a very common occurrence for humans, in part because ER protein folding has an over 80% failure rate per Every hand’s a winner, and every hand’s a loser.
I haven’t read papers about ER stress directly influencing plasmalogen abundance. But I’ve curated papers, including several during this Plasmalogens Week, that demonstrate how oxidative stress reduces plasmalogens.
Here’s my BioScan inflammation / oxidative stress panel:
Back to my peroxisomal function panel: I don’t consider my negative Z-scores (below the population mean) of Total PEs and Total PCs to be actionable. Both of them produced positive Z-scores (above the population mean) of their respective total plasmalogens (Total PLEs +1.3, Total PLCs +0.5). I view Total PEs and Total PCs as pools of raw materials for plasmalogen synthesis that are used when needed.
My July 2025 BioScan shows that my current practices provide adequate plasmalogens as compared with unknown populations. It indicates that to produce adequate plasmalogens, I don’t need ProdromeNeuro and ProdromeGlia plasmalogen precursor supplements that bypass normal peroxisomal function plasmalogen synthesis.
This year’s BioScan was a one-time event. I don’t agree with advocates for constantly tweaking health parameters, or obtaining frequent test results for ‘youthful’ targets, or competing with or conforming to other people’s measurements, or unfounded longevity beliefs. It’s every human’s choice whether or not we take responsibility for our own one precious life. Being overly obsessed about one’s health can be among the many symptoms of what’s ruining a person’s life.
I might use a future version of BioScan along with ProdromeNeuro and ProdromeGlia plasmalogen precursor supplements if I had to recover from an accident or some other health emergency that creates a substantial demand for plasmalogens’ antioxidant activities. But I’d first return to past practices I’ve found to be successful in combating oxidative stress, like increasing the frequency of Nrf2 activation by eating broccoli sprouts twice a day rather than once daily.
Continuing Plasmalogens Week with three 2025 papers, starting with a human study that included plasmalogen biomarkers of non-communicable disease fatigue symptoms:
“This study explored the biological mechanisms underlying fatigue in patients with NCDs using a multi-omics approach. Our findings indicate that distinct metabolic pathways, salivary microbiota, and genetic factors may contribute to different dimensions of fatigue, including general, physical, and mental fatigue.
General fatigue is associated with unsaturated fatty acid biosynthesis, indicating its role in lipid metabolism.
Physical fatigue was associated with plasmalogen synthesis, mitochondrial beta-oxidation of long-chain fatty acids, and selenoamino acid metabolism, suggesting a potential contribution of impaired energy production.
Mental fatigue is associated with homocysteine degradation and catecholamine biosynthesis, which may influence cognitive fatigue.
This exploratory study suggests that fatigue in patients with NCDs may involve disruptions in lipid metabolism, neurotransmitter pathways, microbial composition, and genetic variations. Blood-based biomarkers showed better predictive potential for physical fatigue, whereas salivary-based models were more indicative of mental fatigue.
Although our findings support the role of lipid metabolism, the contribution of plasmalogen synthesis remains underexplored. Further studies are needed to validate these findings and understand their mechanisms of action.”
A human study of metabolic dysfunction-associated steatotic liver disease (MASLD) included investigating plasmalogens:
“In this study, we applied untargeted metabolomic profiling to serum samples from individuals with and without MASLD, classified by the Fatty Liver Index, with the goal of identifying characteristic metabolic signatures and pathways that may underlie disease presence and progression. Individuals in the MASLD group displayed significantly higher levels of ALT, AST, ALP, and GGT, reflecting ongoing hepatic injury, cholestasis, and oxidative stress. However, albumin and bilirubin levels remained within normal limits, indicating early to intermediate disease stages rather than advanced fibrosis or cirrhosis.
A consistent and highly significant lipidomic pattern in the MASLD group is the depletion of plasmalogens and sphingomyelins. Depletion of these lipid classes was identified as a hallmark of insulin resistance as defined by the triglyceride-glucose index. In contrast, phosphatidylcholine, phosphatidylethanolamine, and phosphatidylinositol species were elevated in MASLD, pointing toward broader lipid remodeling events.
Reduced plasmalogen and sphingomyelin levels positions their depletion as a core feature of metabolic dysfunction. Plasmalogens are ether phospholipids with strong antioxidant capacity, and their reduction suggests a loss of protective buffering against oxidative stress, one of the main drivers of MASLD progression. Similarly, sphingomyelin depletion implicates altered membrane dynamics and signaling disturbances, further contributing to metabolic dysfunction.
Depletion of plasmalogens 1-(1-enyl-palmitoyl)-2-oleoyl-GPC (P-16:0/18:1), 1-(1-enyl-palmitoyl)-2-linoleoyl-GPC (P-16:0/18:2), 1-(1-enyl-palmitoyl)-2-palmitoyl-GPC (P-16:0/16:0), 1-(1-enyl-palmitoyl)-2-palmitoleoyl-GPC (P-16:0/16:1), 1-(1-enyl-palmitoyl)-2-oleoyl-GPE (P-16:0/18:1), 1-(1-enyl-palmitoyl)-2-linoleoyl-GPE (P-16:0/18:2), and disruption of the glutamate–gamma-glutamyl pathway stand out as central features of metabolic dysfunction in MASLD, with clear potential to inform biomarker discovery, disease classification, and the design of targeted therapeutic strategies.”
https://www.mdpi.com/2218-1989/15/11/687 “Metabolomic Signatures of MASLD Identified by the Fatty Liver Index Reveal Gamma-Glutamyl Cycle Disruption and Lipid Remodeling”
A rodent study investigated dietary sea squirt (AM) plasmalogen ethanolamine (PlsEtn) extract’s and dietary pig liver (PL) phosphatidyl ethanolamine (PtdEtn) extract’s effects on acetaminophen liver injury:
“We investigated dietary effects of PlsEtn from ascidian on chronic hepatic injury in acetaminophen (APAP)-treated mice. Five-week-old male mice were divided into four groups (n = 12), which were treated with experimental diets for two weeks and then the respective APAP-containing diet for five weeks.
Ingested PlsEtn is digested into lysoPlsEtn and free fatty acid in the small intestine. PlsEtn digests are absorbed and are subsequently resynthesized into PlsEtn preferentially with PUFA.
Acetaminophen is a frequently used analgesic and antipyretic. Approximately 90% of APAP is metabolized by UDP-glucuronosyltransferase and sulfotransferase into glucuronic acid and sulfate conjugates, respectively.
5–9% of APAP is metabolized into the highly reactive intermediate N-acetyl-p-benzoquinone imine (NAPQI). This metabolite is considered a pivotal molecule in APAP-induced hepatotoxicity and is conjugated by glutathione (GSH). Excessive NAPQI levels deplete GSH and covalently bind to cellular proteins, resulting in organelle dysfunction, such as mitochondria dysfunction. These impairments induce oxidative stress, cell malfunctions, and subsequently, cell death, such as ferroptosis and apoptosis.
Mice were treated with continuous APAP consumption to induce oxidative stress and impaired lipid metabolism in the liver. Effects of diets were evaluated based on levels of malondialdehyde (MDA), a marker of lipid oxidation, on fatty acid content, and on expression of apoptosis-related proteins in the liver.
The PlsEtn-rich diet effectively suppressed APAP-induced decrease in body and liver weights of mice. However, this suppressive effect was not observed in mice fed a PtdEtn-rich diet. APAP administration decreased the total fatty acid content in the liver, whereas a PlsEtn-rich diet alleviated this decrease and increased the hepatic content of docosahexaenoic acid (DHA).
Owing to the alkenyl linkage, which exhibits antioxidant properties, PlsEtn was expected to markedly suppress hepatic lipid oxidation. However, its suppressive effect was the same extent as that by PtdEtn. Both PlsEtn and PtdEtn contain an ethanolamine base in their structures, and free ethanolamine and its metabolite choline suppress lipid peroxidation. Dietary PlsEtn and PtdEtn may be metabolized into free ethanolamine and its further metabolites, which may alleviate APAP-induced hepatic lipid oxidation.
Dietary ethanolamine glycerophospholipids (EtnGpls) rich in PlsEtn or PtdEtn suppressed APAP-induced lipid oxidation in the liver. Protein expression results revealed that dietary EtnGpls reduced expression of certain apoptosis-related proteins compared to the APAP group. This reduction was more effective in mice fed the PlsEtn-rich diet than in those on the PtdEtn-rich diet.”
https://www.mdpi.com/2076-3417/15/11/5968 “Dietary Ethanolamine Plasmalogen from Ascidian Alleviates Chronic Hepatic Injury in Mice Treated with Continuous Acetaminophen”
This study neither demonstrated nor provided citations for its dietary plasmalogen recycling statements.
Three more plasmalogen health and disease papers are curated in Part 2.
A 2025 clinical trial with old people compared NRF2 effects of acute exercise with pre- and post-exercise sulforaphane treatment:
“This study tested the hypothesis that combining acute exercise (in vivo stimulus) with ex vivo sulforaphane (SFN) treatment would induce greater NRF2 activation and signaling in older adults compared to either treatment alone. This approach was used to bypass the potential issue of inter-individual variability in metabolism and bioavailability of SFN supplementation through oral consumption and thereby provide more rigorous biological control to establish mechanistic feasibility.
Twenty-five older adults (12 men, 13 women; mean age: 67 ± 5 years) performed 30-min cycling exercise. Blood was drawn before and immediately after exercise to isolate peripheral blood mononuclear cells (PBMCs) and incubate with and without SFN (5 μM) treatment.
Acute exercise induced modest transcriptional changes across the four tested transcripts compared to the robust upregulation elicited by SFN. This disparity was notable given the comparable NRF2/ARE binding activity observed between EX and SFN.
Near-significant trends were observed for EX in heme oxygenase-1 (HO-1), NAD(P)H quinone oxidoreductase 1 (NQO1), and glutathione reductase (GR) (after Bonferroni correction), while glutamate-cysteine ligase catalytic subunit (GCLC) was not induced by EX. In contrast, SFN alone robustly induced expression of NQO1, HO-1, GR, and GCLC.
We had chosen 5 μM as the dose based on pilot data from our laboratory and existing literature from in vitro experiments. However, typically, SFN is not combined with another stimulus.
To test this speculation, we ran a post hoc dose–response experiment where we stimulated PBMCs (n = 5) at six different SFN concentrations ranging from 0 to 20 μM (incubated for 5 h) and analyzed responses across the four genes used in the present study. The dose responses displayed hormetic curves for NQO1, GR, and GCLC, with 5 μM eliciting the peak response, suggesting that the lack of difference between SFN and the combined treatment was due to a ceiling effect of the SFN dose. Interestingly, HO-1 displayed a linear/curvilinear response with the maximal observed response at 20 μM.
In future ex vivo studies, a sulforaphane concentration of 1–2 μM in combination with acute exercise is predicted to enhance the expression of these antioxidant genes in the PBMCs of older adults to a greater extent than either treatment alone. Furthermore, lower SFN plasma concentrations are more likely to be achievable with oral supplementation.
To our knowledge, this is the first trial to measure responses to acute exercise combined with sulforaphane stimulation on NRF2 signaling in older men and women. We did not observe any statistically significant differences in any of our outcome variables between men and women.
Our results demonstrate that combining acute exercise with a sulforaphane stimulus elicits a greater response in nuclear NRF2 activity in older adults. While the response in gene expression did not completely mirror the response in NRF2 activation, it is important to note that NRF2 induces hundreds of cytoprotective genes. The four transcripts we measured are among those most commonly used to represent NRF2 signaling but do not capture the full picture. Full transcriptomics in future studies would address this question.”
I asked two questions, and will follow up with replies:
Did a second experiment test effects of these subjects eating broccoli sprouts prior to acute exercise? The clinical trial’s NCT04848792 Study Overview section indicated that was the researchers’ intent.
What studies have the data that produced this study’s graphical abstract’s younger vs. older NRF2 response graph?